

Coxiella burnetii: A review focusing on infections in German sheep and goat flocks
Berliner und Münchener Tierärztliche Wochenschrift 133
DOI: 10.2376/0005-9366-19030
© Schlütersche Verlagsgesellschaft mbH & Co. KG. 2020
Publiziert: 04/2020
Zusammenfassung
Q-Fieber ist eine weltweit verbreitete Zoonose, die durch Coxiella (C.) burnetii verursacht wird. Dieses Bakterium wird besonders über Geburtsmaterial infizierter kleiner Wiederkäuer in großen Mengen ausgeschieden, aber auch über Milch, Kot und Urin. Der Hauptinfektionsweg für Mensch und Tier ist die Inhalation von erregerhaltigen Aerosolen. Bei der Verbreitung spielen Wind und andere meteorologische Faktoren eine Rolle. Auch Zecken wie Dermacentor spp. und Ixodes ricinus könnten eine Bedeutung bei der Übertragung von C. burnetii haben. Bei Ziegen kann eine Infektion mit C. burnetii vor allem zu Spätaborten führen. Im Gegensatz dazu scheint eine Coxiellen-Infektion bei Schafen zu weitaus geringeren Abortraten als bei Ziegen zu führen. Beim Menschen zeigen 40 % der infizierten Personen grippeähnliche Symptome wie Fieber und Kopfschmerzen. Bis zu 5 % aller Infizierten können eine Chronifizierung entwickeln, die meist mit einer Endokarditis einhergeht. In Deutschland wurden seit dem Jahr 2000 mehrere humane Q-Fieber-Kleinraumepidemien mit bis zu 331 infizierten Personen pro Ausbruch detektiert. Auslöser waren meist lammende Schafe. Aufgrund der Zunahme an Milchziegenhaltungen in Deutschland wird in Zukunft mit vermehrten Fällen auch bei dieser Tierart gerechnet. Der diagnostische Nachweis von Infektionen mit C. burnetii beim Tier erfolgt meist mit dem ELISA und/oder der PCR. Mithilfe der Multiple Loci Variable Number of Tandem Repeat Analysis (MLVA/VNTR) lassen sich Isolate genotypisieren. Das genotypische MLVA-Cluster A wurde besonders bei kleinen Wiederkäuern nachgewiesen, während das Cluster C eher mit Rindern assoziiert ist. Die antibiotische Behandlung infizierter Tiere mit Oxytetracyclinen hat sich nicht bewährt. Vielmehr senkt der Einsatz eines Impfstoffes mit inaktivierten Phase-I-Bakterien das Risiko der Ausscheidung. Aufgrund der hohen Tenazität von C. burnetii sind viele Desinfektionsmittel nicht wirksam.
Summary
Q fever is a worldwide zoonosis caused by Coxiella (C.) burnetii. This bacterium is shed in huge amounts with birth products from infected small ruminants. Shedding also occurs via milk, faeces and urine. The main route of infection for humans and animals is via inhalation of contaminated aerosols. Apart from wind, many other meteorological conditions influence pathogen dissemination. Also, Dermacentor spp. and Ixodes ricinus ticks could play a role in transmission. Infections in goats can lead to abortion during the last trimester of pregnancy. In comparison to goats, C. burnetii seems to cause less abortion in sheep. In humans, 40% of infected persons show flu-like symptoms such as fever and headaches. Up to 5% of all infected people can develop chronic Q fever, which often manifests as endocarditis. Since 2000, several human Q fever epidemics occurred in Germany with up to 331 reported human infections. The source of infection was mostly associated with parturition of sheep. As the number of goats has increased in Germany over the last couple of years, Q fever cases connected to this species will probably increase in the future. For the detection of C. burnetii in animals, ELISA and/or PCR are routinely used. With Multiple Loci Variable Number of Tandem Repeats Analysis (MLVA/VNTR) it is possible to genotype isolates. MLVA-Cluster A was detected in samples from small ruminants, Cluster C is associated with cattle. Treatment with oxytetracycline is ineffective for the control of Q fever in animals, whereas the use of an inactivated Phase I whole cell vaccine reduces shedding and therefore the risk of transmission. Many disinfectants are ineffective against C. burnetii because of its high tenacity.
Einleitung
Q-Fieber ist mit Ausnahme von Neuseeland und der Antarktis eine weltweit verbreitete Zoonose. Der Erreger des Q-Fiebers ist Coxiella (C.) burnetii, ein obligat intrazelluläres, unbewegliches, Gram-negatives Bakterium. In Australien löste der Erreger erstmals 1935 einen Ausbruch bei Schlachthausmitarbeitern aus. Da die Ursache unklar war, wurde diese grippeartige Erkrankung als unklares/fragliches Fieber bzw. „Query (Q) fever“ (Q-Fieber) bezeichnet (Hechemy 2012). Eine Infektion von Tieren mit C. burnetii wird im deutschsprachigen Raum als Coxiellose bezeichnet, aber im internationalen Sprachraum ist die Bezeichnung „Q fever“ auch für Tiere gebräuchlich.
Besonders in den 1990er Jahren kam es immer wieder zu größeren Q-Fieber-Epidemien in Bulgarien (Georgiev et al. 2013, Serbezov et al. 1999). Allein 1993 wurden über 1000 humane Fälle mittels Serologie bestätigt (Martinov 2007). Die Epidemien ließen sich besonders auf Geburten bei Ziegen zurückführen, deren Zahl sich im gleichen Zeitraum mehr als verdoppelt hat (Serbezov et al. 1999). Der weltweit bisher größte humane Q-Fieber-Ausbruch wurde in den Niederlanden registriert. Von 2007 bis 2011 erkrankten dort mehr als 4000 Personen an Q-Fieber (Kampschreur et al. 2013). Allein im Jahre 2009 waren es 2357 Patienten, von denen 459 stationär behandelt werden mussten. Vorerkrankungen führten nach einer Coxiellen-Infektion bei sechs Patienten zum Tod (van der Hoek et al. 2010). Auslöser dieser Epidemie waren Betriebe mit großen Milchziegenherden (durchschnittlich 1000 Tiere) im Süden der Niederlande. Während der Lammung im Frühjahr kam es in infizierten Betrieben zu einer starken Erregerfreisetzung, anfänglich verbunden mit Abortraten von bis zu 60 % pro Betrieb (van den Brom und Vellema 2009). Im Gegensatz dazu wurden Häufungen von Humaninfektionen in Deutschland meist auf lammende Schafe zurückgeführt. Zusätzlich treten Humanerkrankungen mit Q-Fieber gehäuft entlang von Triebwegen auf (Hellenbrand et al. 2001). Die deutsche Ziegenpopulation spielte in den letzten Jahrzehnten eine eher untergeordnete Rolle bei der Übertragung. Obwohl die Prävalenz in Rinderbeständen in einigen deutschen Regionen hoch ist (Böttcher et al. 2011b, Hilbert et al. 2014), sind von Rindern ausgeschiedene Coxiellen als Ursache von Kleinraum-Epidemien von geringerer Bedeutung. Rinder sind vermutlich eher für humane Einzelerkrankungen verantwortlich und diese werden möglicherweise nicht diagnostiziert. Infektionen größerer Personengruppen werden dagegen meist von kleinen Wiederkäuern verursacht. Das gehäufte Auftreten von Krankheitsfällen beim Menschen begünstigt zudem das Erkennen einer Q-Fieber-Erkrankung. Weiterhin könnten physiologische Unterschiede zwischen großen und kleinen Wiederkäuern von Bedeutung sein, wie beispielsweise die unterschiedliche Kotkonsistenz und Ausscheidungsdynamik von C. burnetii. Milchkühe scheiden den Erreger über einen längeren Zeitraum besonders mit der Milch und weniger mit Vaginalsekret und Kot aus (Guatteo et al. 2007, Rodolakis et al. 2007). Bei kleinen Wiederkäuern findet eine Ausscheidung des Erregers hauptsächlich über Kot und Vaginalschleim statt, bei Ziegen auch mit der Milch (Arricau-Bouvery et al. 2003, Berri et al. 2001, Rodolakis et al. 2007). Somit scheint die Umweltkontamination durch infizierte kleine Wiederkäuer größer zu sein als bei Rindern (Arricau-Bouvery et al. 2003). Außerdem könnten die unterschiedlichen Haltungsbedingungen eine Rolle spielen, da Milchkühe in Deutschland meist im Stall und Schafe vor allem im Freien gehalten werden (Ganter et al. 2011).
Der Erreger
Ursprünglich wurde C. burnetii taxonomisch in die Familie der Rickettsiaceae eingegliedert. Analysen des 16S rRNA-Gens und später des kompletten Genoms zeigten jedoch, dass C. burnetii phylogenetisch eng mit Legionella pneumophila, dem Erreger der Legionärskrankheit, verwandt ist (Seshadri et al. 2003, Stein et al. 1993, Weisburg et al. 1989), weshalb das Bakterium innerhalb der Klasse Legionellales in eine eigene Familie Coxiellaceae eingegliedert wurde. Bislang war C. burnetii die einzige Art in der Familie der Coxiellaceae. In den letzten Jahren wurden jedoch noch weitere, neue bzw. Coxiella-ähnliche Bakterien identifiziert, wie beispielsweise C. cheraxi sp. nov. im australischen Flusskrebs (Cherax quadricarinatus), Candidatus C. massiliensis und Candidatus C. mudrowiae in Zecken und Candidatus C. avium in Papageien und einem Tukan (Angelakis et al. 2016, Gottlieb et al. 2015, Shivaprasad et al. 2008, Tan und Owens 2000). C. burnetii selbst wurde in einer Vielzahl verschiedener Spezies der Arthropoden, Vögel und Säugetiere nachgewiesen (Woldehiwet 2004).
C. burnetii existiert in zwei verschiedenen Formen: den sogenannten small-cell-variant (SCV) und large-cell-variant (LCV) (McCaul und Williams 1981). Die SCV sind die stoffwechselphysiologisch dormante und nicht replikative Form. Sie besitzen eine hohe Tenazität mit Resistenz gegenüber Austrocknung, Hitze, UV-Licht und zahlreichen Desinfektionsmitteln (Eldin et al. 2017). Das Bakterium kann bis zu 32 Monate in Rohmilch nachgewiesen werden und bis zu zwei Jahre bei 4 ºC im Staub und Wolle überleben (Angelakis und Raoult 2010, Schliesser 1991). Im Gegensatz dazu, konnten in einer neueren Studie von Álvarez-Alonso et al. (2018) vermehrungsfähige Coxiellen aus Stallstaubproben nur innerhalb von zwei Monaten nach der letzten Lammung mittels Inokulation in Mäuse mit anschließender Kultivierung in Vero-Zellen festgestellt werden. Diese Diskrepanz bedarf zukünftig weiterer Abklärungen. Die LCV sind die metabolisch aktive Form und werden im Phagolysosom der Wirtszelle nachgewiesen (Coleman et al. 2004, Minnick und Raghavan 2012).
Eine weitere Besonderheit von C. burnetii ist die phasenspezifische Antigenvariation. Wie bei Enterobakterien ist das Lipopolysaccharid (LPS) das dominierende Oberflächenantigen auf der äußeren Membran von C. burnetii. Bakterien, die ein langkettiges, glattes LPS mit komplettem O-spezifischen Polysaccharid exprimieren, liegen als Phase I (Ph I) vor. Sie sind virulent und können aus Wirten wie Mensch, Tier und Zecke isoliert werden. Durch häufiges Passagieren in immuninkompetenten Wirten, z. B. embryonierten Hühnereiern, verkürzt sich das LPS zunehmend. Der Verlust des LPS geht mit einer Verringerung der Virulenz einher. Bakterien mit einem stark verkürzten, rauen LPS sind gering bis avirulent und werden als Coxiellen in Phase II (Ph II) bezeichnet (Hotta et al. 2004).
Top Job:
Übertragung und Infektion
Tierhaltungen
C. burnetii wird über Geburtsmaterial (Plazenta, Amnionflüssigkeit) von infizierten Schafen und Ziegen während der Geburt oder eines Abortes in großen Mengen ausgeschieden. In 1 g Plazentagewebe können bis zu 109 C. burnetii-Erreger enthalten sein (Babudieri, zitiert nach Fournier et al. 1998). C. burnetii vermehrt sich insbesondere in den Trophoblasten der Plazenta, wobei die Vermehrungsrate zur Geburt hin zunimmt (Ben Amara et al. 2010, Roest et al. 2012). Warum C. burnetii speziell in Trophoblasten ideale Bedingungen für eine Vermehrung findet, ist noch nicht vollständig geklärt. Die Fähigkeit von C. burnetii, die Transkription in Trophoblasten umzuprogrammieren, scheint dabei eine wesentliche Rolle zu spielen (Ben Amara et al. 2010). Eine Ausscheidung findet auch über Milch, Kot und Urin statt. Aufgrund der hohen Tenazität des Erregers gegenüber Umwelteinflüssen, lässt sich die DNS von C. burnetii ubiquitär nachweisen (Kersh et al. 2010). Größere Mengen werden jedoch besonders in Tierhaltungen von Wiederkäuern gefunden (Carrie et al. 2019, Kersh et al. 2010). Der Erreger des Q-Fiebers ist hoch kontagiös. Der Hauptinfektionsweg für Mensch und Tier ist die Inhalation erregerhaltiger Aerosole (Brooke et al. 2013, Todkill et al. 2018). Die aerogene Infektion nach Verbreitung des Erregers mit dem Wind hat in verschiedenen europäischen Ländern zu urbanen Ausbrüchen innerhalb eines begrenzten Umfelds (Kleinraumepidemien) geführt (Gilsdorf et al. 2008, Hawker et al. 1998, Tissot-Dupont et al. 1999). Ein besonderes Infektionsrisiko besteht innerhalb einer 5 bis 10 km Zone um das Ausbruchsgeschehen herum, wobei das größte Risiko innerhalb eines Radius von 2 bis 4 km liegt (Clark und Soares Magalhães 2018). Jedoch wurde auch von einer Übertragung von bis zu 18 km Entfernung berichtet (Hawker et al. 1998). Von einer anderen Arbeitsgruppe konnte im Gegensatz dazu C. burnetii nur in einem Umkreis von 50 Metern um einem positiven Ziegenbetrieb nachgewiesen werden (Kersh et al. 2013). Bei einer Kleinraumepidemie in Jena im Jahre 2005 erkrankten Menschen, die in einem Umkreis von 400 Metern um das Ausbruchsgeschehen wohnten (Gilsdorf et al. 2008). Zusätzlich kann C. burnetii auch durch frischen Schaf- und Ziegenmist von infizierten Betrieben verbreitet werden. Dieser ist besonders durch Fruchtwasser und nicht entfernte Nachgeburten massenhaft mit C. burnetii kontaminiert. Die Ausbringung auf landwirtschaftlichen Flächen, vor allem bei windigem Wetter, hat in der Vergangenheit immer wieder zu humanen Q-Fieber-Erkrankungen und Ausbrüchen geführt (Arricau-Bouvery und Rodolakis 2005, King et al. 2011, Reintjes et al. 2000). Obwohl es Hinweise gibt, dass die Mistausbringung im Falle des Ausbruchs in den Niederlanden eine untergeordnete Rolle spielte (van den Brom et al. 2015a), wurde später in einem Konsensuspapier eine dreimonatige Mistlagerung unter Folie und der Transport des Mistmaterials nur an windstillen Tagen empfohlen (Plummer et al. 2018). Neben dem Wind spielen auch andere meteorologische Faktoren eine Rolle. So begünstigt z. B. trockenes Wetter die Ausbreitung von Coxiellen (Tissot-Dupont et al. 2004), Regenfälle können dagegen die Ausbreitung und damit das Risiko für humane Infektionen verringern (Gilsdorf et al. 2008).
Rohmilch
Ob der Genuss von mit C. burnetii kontaminierter Rohmilch und Rohmilchprodukten beim Menschen zur Erkrankung führt, wird kontrovers diskutiert (Cerf und Condron 2006, Eldin et al. 2013, Raoult et al. 2000). Bereits in den 1960er Jahren konnte in einer Studie an Gefängnisinsassen gezeigt werden, dass der Konsum von mit C. burnetii kontaminierter Rohmilch zur Serokonversion, aber nicht zur Erkrankung führt (Benson et al. 1963). Eine solche Studie kann selbstverständlich aus ethischen Gründen nicht wiederholt werden. Es wird vermutet, dass die aerogene Infektion aufgrund der höheren Anzahl von Makrophagen in der Lunge effektiver ist als eine orale Infektion (Gale et al. 2015). Dennoch scheint es in wenigen Fällen beim regelmäßigen Konsum von erregerhaltiger Rohmilch zu Erkrankungen mit Fieber und starken Kopfschmerzen zu kommen (Fishbein und Raoult 1992, Signs et al. 2012). Dupont et al. (1992) vermuten auch einen Zusammenhang zwischen Rohmilchkonsum und C. burnetii-assoziierter Hepatitis. Dieses Erkrankungsbild ist besonders in Frankreich vorherrschend (Dupont et al. 1992, Melenotte et al. 2018). Aktuell wird das Risiko, durch den Konsum von Rohmilch und Rohmilchprodukten an Q-Fieber zu erkranken, als gering, aber nicht vernachlässigbar eingestuft (Gale et al. 2015, Pexara et al. 2018). Besonders vor dem Hintergrund, dass Rohmilchkäse lebende Coxiellen beinhalten kann, sollte Milch aus infizierten Herden grundsätzlich einer Wärmebehandlung unterzogen werden (Barandika et al. 2019, BfR 2010). Durch den Prozess der Pasteurisierung (71,66 °C für 15 Sekunden) wird der Erreger in der Milch inaktiviert (EFSA 2010).
Zecken
Zecken sind Überträger zahlreicher Zoonose-Erreger. Die Rolle der Zecken als Reservoir und Überträger von C. burnetii ist noch nicht eindeutig geklärt. Weltweit wurde der Erreger in unterschiedlichen Zeckenarten nachgewiesen (Duron et al. 2015). In Deutschland ist Ixodes (I.) ricinus die mit Abstand meist verbreitete und häufigste Zeckenart (Rubel et al. 2014). Obwohl Dermacentor (D.) marginatus, auch Schafzecke genannt, nicht flächendeckend im gesamten Bundesgebiet vorkommt (Rubel et al. 2014, Walter et al. 2016), gilt diese als Vektor für C. burnetii (Sting et al. 2004). Allerdings konnte C. burnetii nur in einer von 1066 untersuchten Zecken der Spezies D. marginatus mittels PCR nachgewiesen werden, obwohl die Zecken aus einem Q-Fieber endemischen Gebiet stammten (Sting et al. 2004). Wenige Jahre später konnte im gleichen Untersuchungsgebiet weder in D. marginatus noch in D. reticulatus Coxiellen-DNS detektiert werden (Pluta et al. 2010). Nach der großen Q-Fieber-Epidemie in den Niederlanden wurden 1891 DNS-Lysate von Zecken mittels multiplex qPCR getestet. C. burnetii wurde in keiner Probe gefunden (Sprong et al. 2012). Jedoch konnten Hildebrandt et al. (2011) C. burnetii in einem thüringischen Endemiegebiet bei 1,9 % der untersuchen I. ricinus nachweisen. Insgesamt dürften daher sowohl Dermacentor spp. als auch I. ricinus epidemiologisch eher eine untergeordnete Rolle bei der Übertragung von C. burnetii in Deutschland spielen (Kimmig 2010). Neuere Studien zeigen, dass etwa 75 % der Zeckenarten Coxiella-ähnliche Bakterien beherbergen, dies könnte in der Vergangenheit zu falsch positiven Erregernachweisen beigetragen haben (Duron 2015, Jourdain et al. 2015).
Die Infektion mit C. burnetii durch Zeckenstich ist bisher ebenfalls nicht eindeutig geklärt (Duron et al. 2015, González-Barrio et al. 2016). Die Übertragung mittels Zeckenkot spielt für die Infektion von Wirten wahrscheinlich eine größere Bedeutung als der Zeckenstich (Liebisch 1977). Coxiellen vermehren sich in Darmzellen der Zecken sehr stark und werden mit dem Kot ausgeschieden (Angelakis und Raoult 2010, Liebisch 1977, Špitalská und Kocianová 2003). Der erregerhaltige Zeckenkot (bis zu 1012 Coxiellen/g) verklebt die Wolle und die darin enthaltenen Coxiellen können bis zu 2 Jahre infektiös bleiben (Schliesser 1991). Schulz et al. (2005) konnten im Luftraum der Umgebung während der Schur Coxiellen-DNS nachweisen. Humane Infektionen durch Inhalation des kontaminierten Staubs sind somit während der Schur möglich (Angelakis und Raoult 2010). Deshalb sollten auch bei der Schur entsprechende Vorsichtsmaßnahmen getroffen werden, um eine Infektion zu verhindern.
Bakteriämie und Immunantwort
Bei Mensch und Tier erfolgt die Coxiellen-Infektion vor allem aerogen mittels kontaminierter Stäube und/oder Tröpfchen (Kimmig 2010). Möglicherweise können hier die Alveolar-Makrophagen eine entscheidende Rolle spielen, da beim Einatmen kontaminierter Stäube und/oder Aerosole der erste Erregerkontakt mit dem Wirt in der Lunge erfolgt (Khavkin und Tabibzadeh 1988). Gefolgt von dieser ersten Erregeraufnahme und Vermehrung sind eine hämatogene Streuung und spezifische Organmanifestation (hervorzuheben sind hier Lunge, Gehirn, Leber sowie Fortpflanzungsorgane (Kimmig 2010, Schüle 2008)) als Folge der Bakteriämie zu beobachten. Die betroffenen Organe sind mit Blick auf humane Infektionen bisher noch nicht vollständig erfasst. Während der systemischen Ausbreitung sind auf zellulärer Ebene vor allem Alveolar-Makrophagen und Kupffer’sche Sternzellen in der Leber von Infektionen betroffen. Im weiteren Verlauf können beim Menschen eine Pneumonie oder eine granulomatöse Hepatitis beobachtet werden (Kimmig 2010). C. burnetii wird während der Bakteriämie und darüber hinaus über verschiedene Körperflüssigkeiten ausgeschieden (Rousset et al. 2009a, Schüle 2008). Der Tropismus des Erregers betrifft bei Wiederkäuern besonders Epithelzellen der Kotyledonen und resultiert in einer hohen Bakterienkonzentration in der Amnionflüssigkeit, der Plazenta sowie dem Lochialsekret. Die hohe Erregerlast kann Frühgeburten oder Aborte bewirken und erhöht das postnatale Infektionsrisiko. Primäre Wirtszellen von C. burnetii sind Makrophagen. Infektionen von Trophoblasten, Endothel- und Epithelzellen sowie von Fibroblasten wurden nachgewiesen (Maurin und Raoult 1999, Voth und Heinzen 2007). Als intrazelluläres Pathogen ist es Coxiellen möglich, sich der Antikörper-Opsonierung sowie der Eliminierung über das Komplementsystem zu entziehen (Bonkowski 2016). Makrophagen nehmen Bakterien über den Mechanismus der Phagozytose in intrazelluläre Phagosomen auf. Dieses vakuoläre Zellkompartiment reift durch Fusion mit Endosomen und Lysosomen zu einem degradationsaktiven Phagolysosom, in dem die aufgenommenen Bakterien im Allgemeinen abgetötet werden können (Flannagan et al. 2009). Coxiellen haben Mechanismen entwickelt, in den Phagolysosomen zu überleben und sich zu vermehren (Howe et al. 2010). Neben den Makrophagen sind auch dendritische Zellen (DCs) Wirtszellen für C. burnetii (Shannon et al. 2005) (Abb. 1). Mithilfe der toll-like receptors (TLRs) bzw. der damage-associated molecular pattern receptors (DAMP-R) verfügen DCs über wichtige Sensoren des angeborenen Immunsystems und modulieren nachgeordnete Immunantworten (Mosser und Edwards 2008). Ein zentraler Mechanismus angeborener Immunität ist die Stimulation von Pathogenmuster-Rezeptoren (pattern recognition receptors, PRRs) und DAMP-Rs der betreffenden Immunzellen. Als relevante Aufnahmerezeptoren dienen Integrine (αvβ3, für LPS-Phasentyp I und II) sowie der Komplementrezeptor 3 (CR3, nur LPS-Phasentyp II) (Capo et al. 1999). Nach dem Eindringen in die Wirtszelle nutzen Coxiellen einen für intrazellulären Abbau und zellautonome Immunabwehr vorgesehenen autophagischen Weg für ihre intrazelluläre Vermehrung (Colombo et al. 2006). Autophagie ist ein evolutionär konservierter Prozess aller eukaryotischen Zellen, der normalerweise dazu dient, nicht mehr benötigte oder defekte Zellbestandteile abzubauen und/oder zu recyceln. Darüber hinaus kann die Autophagie nach Infektion intrazelluläre Pathogene mittels xenophagischer Degradation, bei der die Autophagie gegen Krankheitserreger gerichtet ist, beseitigen. Nach der Aufnahme von Coxiellen durch die Wirtszelle entlang des Phagozytosewegs kommt es zur Ausbildung einer parasitophoren Vakuole (PV). Im weiteren Verlauf fusioniert diese mit autophagischen Kompartimenten, eine wichtige Voraussetzung für die erfolgreiche intrazelluläre Anpassung an die Wirtszelle. Interessanterweise entgeht C. burnetii hierbei der Degradation und kann sich ungehindert in den Autophagiekompartimenten vermehren (Voth et al. 2007). Anhand unterschiedlicher Autophagiemarker ließ sich der intrazelluläre Transportprozess von Coxiellen im Detail nachvollziehen (Colombo et al. 2006, Lührmann et al. 2017, Schulze-Luehrmann et al. 2016, Voth et al. 2007). Durch ihr Typ-IVB-Sekretionssystem (T4BSS) sind Coxiellen ideal an eine intrazelluläre Lebensweise angepasst und können Virulenzproteine, sogenannte Effektorproteine, in die Wirtszelle exportieren (van Schaik et al. 2013) (im nachfolgenden Abschnitt näher beschrieben). Funktionell sind bakterielle Effektoren (Lührmann et al. 2017) verantwortlich für die Ausbildung replikativer PVs (Beare et al. 2011, Carey et al. 2011), die Inhibition von Xenophagie (Newton et al. 2014, Winchell et al. 2014), die Modulation der Reifung der PV (Larson et al. 2013, Latomanski et al. 2016, Martinez et al. 2016), die Inhibition des Zelltodes (Beare et al. 2011, Bisle et al. 2016, Cunha et al. 2015, Lührmann et al. 2010), die Remodellierung des Zytoskeletts (Weber et al. 2016) und die Unterwanderung der Immunantwort (Clemente et al. 2018). Bei Monozyten/Makrophagen unterscheidet man zwischen klassischer M1- sowie alternativer M2-Polarisierung. Während antigen-präsentierende proinflammatorische M1-Monozyten/Makrophagen eine intrazelluläre Bakterienvermehrung kontrollieren, wird diese bei antiinflammatorischen M2-Monozyten/Makrophagen begünstigt (Benoit et al. 2008b). Die Interaktion von Monozyten/Makrophagen mit C. burnetii führt zu unterschiedlicher Polarisierung. So induzieren Coxiellen in ruhenden Monozyten, in denen C. burnetii überlebt, sich aber nicht vermehrt, eine M1-Polarisierung (Ghigo et al. 2004). Hingegen führt in Makrophagen, in denen C. burnetii repliziert, die entsprechende Infektion zu einer M2-Polarisierung mit Induktion antiinflammatorischer Zytokine (Interleukin (IL)-10, transforming growth factor (TGF)-β und IL-1Ra), welche sowohl die Vermehrung als auch die Persistenz des Erregers unterstützen (Benoit et al. 2008a). Des Weiteren zeigte sich, dass es zu keiner Induktion einer für die M1-Polarisierung charakteristischen Antigenpräsentation infizierter Makrophagen kommt (Sobotta et al. 2016). Da die Stimulierung des Immunsystems über konservative PRRs wie extrazelluläre TLRs (Ammerdorffer et al. 2015, Bradley et al. 2016, Ramstead et al. 2016, Zamboni et al. 2004) und intrazelluläre NLRs (NOD-like-Rezeptoren) (Ammerdorffer et al. 2015) vermittelt wird, stehen diese im Fokus aktueller Studien zu C. burnetii. Beim Menschen sind einige genetische PRR-Varianten mit einer höheren Wahrscheinlichkeit assoziiert, eine chronische Verlaufsform des Q-Fiebers zu entwickeln (Schoffelen et al. 2015). In diesem Zusammenhang ist interessant, dass das LPS virulenter Coxiellen-Stämme bei infizierten Makrophagen möglicherweise als eine Art TLR4-Antagonist fungiert (Zamboni et al. 2004). Obwohl der TLR4 für die späte Kontrolle der C. burnetii-Infektion entbehrlich zu sein scheint, kontrolliert es die frühen Ereignisse der Infektion, einschließlich Makrophagenphagozytose, Granulombildung und Zytokinproduktion (Honstettre et al. 2004).
Bei der Infektion von DCs mit C. burnetii zeigt sich weder eine Induktion funktionaler DC-Reifung noch eine entsprechende Zytokinproduktion (Shannon et al. 2005). Obschon der verantwortliche molekulare Mechanismus noch unklar ist, wird angenommen, dass hier das Coxiellen-LPS als Virulenzfaktor eine funktionelle DC-Stimulation blockiert (Shannon et al. 2005). Es wird postuliert, dass es möglicherweise den Zugang bakterieller Oberflächenmoleküle zu den entsprechenden TLRs infizierter DCs auf sterische Weise beeinträchtigt. Das Ausbleiben der Reifung infizierter DCs führt vermutlich zur Toleranz gegenüber C. burnetii-Antigenen und kann so für die Etablierung und/oder Manifestation persistenter Infektionen verantwortlich sein (Steinman und Nussenzweig 2002, Verhasselt et al. 2004). Eine erfolgreiche Präsentation von Antigenen mithilfe aktivierter Antigen-präsentierender Zellen (APCs) führt zur Einleitung der adaptiven Immunantwort, die eine entscheidende Rolle bei der Coxiellen-Abwehr spielt. So zeigte sich im Maussystem mit fehlender T- und B-Zellimmunität eine ungehinderte C. burnetii-Infektion (Andoh et al. 2007, Islam et al. 2013, Kishimoto et al. 1978). T-Zell-depletierte Versuchstiere, welche nur eine antikörperbasierte Immunantwort besitzen, können eine Infektion ebenfalls nicht eliminieren (Read et al. 2010). Die für die Bekämpfung von Coxiellen erforderliche TH1-vermittelte zelluläre Immunität basiert größtenteils auf der Ausschüttung proinflammatorischer Zytokine, wie Interferon-γ (IFN-γ). Eine eingeschränkte oder ausbleibende IFN-γ-Produktion der T-Zellen äußert sich in einer erhöhten Mortalitätsrate infizierter Versuchstiere (Andoh et al. 2007). Obschon die IFN-γ-Aktivierung infizierter Zellen die C. burnetii-Replikation vor allem durch die Produktion reaktiver Sauerstoff- (ROS) und Stickstoffspezies (RNO) hemmt (van Schaik et al. 2013), scheinen die Bakterien in der Lage zu sein, die ROS-Produktion über die Sekretion einer sauren Phosphatase zu verhindern (Hill und Samuel 2011). Es wird vermutet, dass diese in Wirtszellen die Bildung des NADPH-Oxidase-Komplexes verhindert (Siemsen et al. 2009). Die Produktion von RNO erfordert die Neusynthese der induzierbaren Stickoxidsynthase iNOS (Vila-del Sol et al. 2007), wie sie nach TLR-Stimulation und nachfolgender Induktion proinflammatorischer Zytokine zu beobachten ist. Jedoch führt aufgrund des Ausbleibens dieser Stimulation (Benoit et al. 2008a) die Coxiellen-Infektion von Makrophagen nicht zu einer entsprechenden RNO-Synthese. Interessanterweise zeigen in diesem Zusammenhang kürzlich veröffentliche Studien zu bovinen und humanen Makrophagen, die mit C. burnetii-Stämmen unterschiedlicher Genotypen infiziert wurden, dass die Hochregulation von Aktivierungsmarkern (CD40 und CD80) und die proinflammatorische Zytokinantwort infizierter Makrophagen, durch den jeweiligen Genotyp des C. burnetii-Stamms bestimmt wird (Sobotta et al. 2016).
Coxiellen-Wirtszell-Interaktionen greifen demnach mannigfaltig in die Immunität ein. Coxiellen sind in der Lage, der zelleigenen Abwehr zu entgehen und Auto-/Xenophagie für sich zu nutzen. Sie manipulieren zudem gezielt die gegen sie gerichtete Immunantwort durch Subversion. Weitere intensive Studien sind notwendig, um das immunologische Verständnis von Infektionen mit C. burnetii zu verbessern und geeignete Immunprophylaxen und Therapien zu entwickeln.
Apoptose
Als obligat intrazelluläres Bakterium kann sich C. burnetii unter natürlichen Bedingungen nur innerhalb infizierter Zellen vermehren. Wie bereits oben beschrieben, ist das Bakterium in der Lage dem intrazellulären Abbau durch Makrophagen zu entgehen und eine entsprechende PV auszubilden (Lührmann et al. 2017). Eine mögliche weitere Abwehrstrategie infizierter Makrophagen in dieser Situation ist die Induktion der Apoptose, wodurch infizierte Zellen durch eine Art zelluläre „Selbsttötung“ eliminiert werden, ohne eine Entzündungsreaktion des adaptiven Immunsystems auszulösen. C. burnetii besitzt jedoch wiederum die Fähigkeit diese entscheidende Wirtszellabwehr zu hemmen (Lührmann und Roy 2007, Voth et al. 2007). Dies ist für das sich langsam replizierende intrazelluläre Bakterium wesentlich, um seine intrazelluläre replikative Nische zu erhalten (Friedrich et al. 2017). Die genauen Mechanismen, mit denen C. burnetii seine Wirtszelle am Leben erhält, sind bisher nicht vollständig geklärt. Allerdings ist klar, dass für diese Aktivität das T4BSS und die von ihm exportierten Virulenz-Faktoren essenziell sind (Beare et al. 2011). Bislang wurden ca. 150 dieser bakteriellen Effektor-Proteine identifiziert. So hemmen AnkG, CaeA und CaeB die Apoptose und IcaA die Pyroptose, ein proinflammatorisches Todesprogramm (Bisle et al. 2016, Cunha et al. 2015, Eckart et al. 2014, Klingenbeck et al. 2013, Lührmann et al. 2010, Schäfer et al. 2017). Allerdings wird angenommen, dass es noch weitere bakterielle Faktoren gibt, die den Zelltod infizierter Wirtszellen verhindern (Martinez et al. 2014) oder induzieren (Schoenlaub et al. 2016, Zhang et al. 2012). Während eine frühe Hemmung des Zelltodes wichtig für die intrazelluläre Vermehrung von C. burnetii ist, könnte die Induktion des Zelltodes zu einem späteren Zeitpunkt der Zellinfektion eine entscheidende Rolle bei der Dissemination der Bakterien im Wirtsorganismus spielen.
Die Fähigkeit von C. burnetii intrazellulär in Makrophagen zu replizieren hängt maßgebend von der Polarisierung des Makrophagen ab. So repliziert C. burnetii in atypischen M2-Makrophagen, während sie in M1-Makrophagen kontrolliert bzw. abgetötet werden (Benoit et al. 2008a, Fernandes et al. 2016, Meghari et al. 2007, Mehraj et al. 2013). Ob C. burnetii die Polarisierung der Makrophagen beeinflusst, ist bislang nicht bekannt. Allerdings konnte nachgewiesen werden, dass Makrophagen durch den Kontakt mit apoptotischen Lymphozyten in Richtung M2 aktiviert werden (Benoit et al. 2008c). Dies könnte auch erklären, weshalb Patienten mit Valvulopathien ein erhöhtes Risiko aufweisen eine Endokarditis zu entwickeln. Diese Patienten weisen mehr apoptotische Leukozyten im Blut auf, was dann zur M2-Aktivierung von Makrophagen und somit zur optimalen Vermehrung von C. burnetii führt (Benoit et al. 2008c).
Klinik bei Ziegen und Schafen
Bei Ziegen kann eine Infektion mit C. burnetii zu Spätaborten und zur Geburt lebensschwacher Lämmer führen (Achard und Rodolakis 2017). Insgesamt können über 90 % der Ziegen in einer Herde davon betroffen sein (Palmer et al. 1983). Meistens werden jedoch Abortraten von 5 bis 20 % beobachtet (Palmer et al. 1983, van den Brom und Vellema 2009). Ziegen, die aufgrund einer C. burnetii-Infektion abortieren, scheinen im Anschluss eher eine Metritis zu entwickeln (van den Brom et al. 2015c). Bezüglich der Intensität der Erregerausscheidung besteht kein Unterschied zwischen Abort und physiologischer Geburt gesunder Lämmer (Roest et al. 2012, Rousset et al. 2009a). Bei Schafen ist das klinische Bild der Coxiellose nicht eindeutig geklärt (Agerholm 2013). Es kann ebenfalls zu einem Spätabort und der Geburt lebensschwacher Lämmer kommen (Berri et al. 2005, Hazlett et al. 2013, Zeman et al. 1989). Infizierte Schafe können aber genauso gesunde Lämmer auf die Welt bringen (Böttcher et al. 2019, Hamann et al. 2009, Welsh et al. 1951). Unter welchen Bedingungen es zum Spätabort oder zur Geburt lebensschwacher Lämmer kommt, bleibt unklar. Grundsätzlich liegt die durch eine C. burnetii-Infektion verursachte Abortrate bei Schafen niedriger als bei Ziegen (Agerholm 2013). Beispielsweise wurde von einer durchschnittlichen Abortrate bei Schafen von 5 % berichtet (van den Brom und Vellema 2009). In den darauffolgenden Lammungen kommt es bei Schafen in der Regel zu keinem Abort mehr (Berri et al. 2002). Bei einem Nachweis von mindestens 104 Erregern in der Plazenta oder im Vaginaltupfer gilt C. burnetii als Verursacher des Abortes (Bru et al. 2013, Hazlett et al. 2013). Derartige und höhere Erregermengen werden auch bei Normalgeburten vorgefunden (Böttcher et al. 2019). Die abortieren Feten sehen unverändert und in der Regel frisch aus, nur gelegentlich sind sie autolytisch (Moeller 2012, van den Brom et al. 2015c). Vereinzelt lässt sich makroskopisch eine Plazentitis mit bräunlich-eitrigen Belägen sowie interkotyledonalen Verdickungen erkennen (Moeller 2012, van den Brom et al. 2015c). Histologisch lassen sich in den interkotyledonalen Trophoblasten sowie an der Basis der Chorionzotten milde mononukleäre Infiltrationen bis hin zu starker Nekrose und purulente Exsudation finden (van den Brom et al. 2015c). Darüber hinaus treten auch Mischinfektionen mit anderen Aborterregern wie Chlamydia (Chl.) abortus und Toxoplasma gondii auf (Berri et al. 2001, Eibach et al. 2013, Hazlett et al. 2013). Deshalb stellt sich besonders bei Schafen die Frage nach der primären Ursache des Abortes, zudem wenig über den Effekt einer Koinfektion der beiden intrazellulären Erreger C. burnetii und Chl. abortus bei kleinen Wiederkäuern bekannt ist.
Vor dem Lammen bzw. dem Abort wird C. burnetii nicht über Vaginalsekret, aber über Kot ausgeschieden (Arricau-Bouvery et al. 2003, Roest et al. 2012). Bei der Ausscheidungsintensität von C. burnetii über die Milch, den Kot und den Vaginalschleim gibt es Unterschiede zwischen Ziegen und Schafen. Während Ziegen den Erreger vor allem auch über die Milch ausscheiden, scheiden Schafe Coxiellen eher über Vaginalschleim und Kot aus (Rodolakis et al. 2007). Über die Ausscheidungsdauer gibt es unterschiedliche Angaben. Arricau-Bouvery et al. (2003) konnten post partum bei experimentell infizierten Ziegen mittels PCR C. burnetii bis zu 14 Tage in Vaginalproben, 20 Tage im Kot und 52 Tage in der Milch nachweisen. Dagegen konnten Roest et al. (2012) bei Ziegen post partum C. burnetii knapp 100 Tage in Vaginaltupfern und im Kot nachweisen, aber nur 38 Tage in der Milch. Bei Schafen gelang es, C. burnetii in Vaginaltupfern bis zu 71 Tage und in Kot sowie Milch bis zu acht Tage nach der Geburt nachzuweisen (Berri et al. 2001). Weiterhin können kleine Wiederkäuer bei der zweiten Lammung den Erreger bis zu vier Monate lang ausscheiden, wenn keine Bekämpfungsmaßnahmen getroffen werden (Astobiza et al. 2011a, Berri et al. 2007). Diese Differenzen bezüglich der Ausscheidungsdauer könnten auch an der unterschiedlichen Sensitivität der verschiedenen Nachweismethoden in den jeweiligen Studien liegen. Auch Kontaminationen während der Probenentnahme in der mit Coxiellen hoch kontaminierten Umwelt können zu falsch-positiven Ergebnissen führen (Plummer et al. 2018, Roest et al. 2012). Bei Milchrindern sind intermittierende Ausscheider oder sogar Dauerausscheider beschrieben (Böttcher et al. 2013, Lucchese et al. 2015). Diese scheiden C. burnetii über einen längeren Zeitraum mit der Milch aus. Van den Brom et al. (2013) konnten vermutlich chronische Milchausscheider auch bei Ziegen nachweisen. Bei Schafen wurde dies bislang noch nicht beschrieben.
Epidemiologische Situation in Deutschland (2000–2018)
Die Schafhaltung in Deutschland variiert sehr stark. Im Ländervergleich haben Schleswig-Holstein (20,77 Schafe/km2), Thüringen (20,66 Schafe/km2) und Baden-Württemberg (17,20 Schafe/km2) die meisten Schafe pro km2 landwirtschaftlich genutzter Fläche, gefolgt von Hessen (16,64 Schafe/km2), Rheinland-Pfalz (11,33 Schafe/km2) und Nordrhein-Westfalen (11,07 Schafe/km2) (Abb. 2). Demgegenüber werden die meisten Ziegen pro km2 landwirtschaftlicher genutzter Fläche in Baden-Württemberg (2,10 Ziegen/km2), Thüringen (1,61 Ziegen/km2), Hessen (1,22 Ziegen/km2) und Bayern (1,21 Ziegen/km2) gehalten. In den anderen Bundesländern spielt die Ziegenhaltung eine untergeordnete Rolle (Abb. 3). Die Anzahl der Schaf- und Ziegenbetriebe pro km2 landwirtschaftlich genutzter Fläche ist in Baden-Württemberg (0,3685 Betriebe/km2), Hessen (0,2834 Betriebe/km2) und Bayern (0,2618 Betriebe/km2) am höchsten. In den nördlichen Bundesländern ist die Anzahl an Schaf- und Ziegenbetrieben am niedrigsten, wobei Nordrhein-Westfalen (0,2080 Betriebe/km2) und Schleswig-Holstein (0,1996 Betriebe/km2) die Ausnahme darstellen (Abb. 4). Betrachtet man die durchschnittliche Anzahl von kleinen Wiederkäuern pro Betrieb ergibt sich ein umgekehrtes Bild. Besonders die Bundesländer Sachsen-Anhalt (durchschnittlich 197 kleine Wiederkäuer/Betrieb), Brandenburg (durchschnittlich 155 kleine Wiederkäuer/Betrieb) und Mecklenburg-Vorpommern (durchschnittlich 139 kleine Wiederkäuer/Betrieb) haben zwar wenige Schaf- und Ziegenbetriebe pro km2 landwirtschaftlicher genutzter Fläche, jedoch halten die wenigen Betriebe im Durchschnitt sehr viele Tiere. Mit Abstand die höchste durchschnittliche Tierzahl pro Betrieb (im Durchschnitt 241 kleine Wiederkäuer/Betrieb) hat Thüringen, während in Bayern eine Vielzahl an kleinen Schaf- und Ziegenhaltungen existieren (im Durchschnitt 62 kleine Wiederkäuer/Betrieb; Abb. 5). Insgesamt ist Baden-Württemberg das Bundesland mit vielen Schafen und Ziegen sowie mit den meisten Schaf- und Ziegenbetrieben pro km2 landwirtschaftlich genutzter Fläche. Dabei handelt es sich um Betriebe mit vergleichsweise mittlerer Tierzahl (im Durchschnitt 90 kleine Wiederkäuer/Betrieb).
Unabhängig von Tierzahl, Anzahl der Betriebe und durchschnittlicher Herdengröße lässt sich die Schafhaltung in Deutschland auch nach gehaltenen Schafrassen und dem damit verbundenem Reproduktionsverhalten in zwei Hälften einteilen. In Süddeutschland (Baden-Württemberg, Bayern, Hessen, Rheinland-Pfalz, Saarland, Sachsen, Thüringen) werden vor allem Merinorassen gehalten, die asaisonal sind und somit ganzjährig lammen (von Korn 2001). Im Gegensatz dazu werden in den nördlichen Bundesländern (Berlin, Brandenburg, Bremen, Hamburg, Mecklenburg-Vorpommern, Niedersachsen, Nordrhein-Westfalen, Sachsen-Anhalt, Schleswig-Holstein) bevorzugt Fleischschafrassen gehalten, die saisonal sind und somit vor allem von Februar bis April lammen (von Korn 2001).
Der Nachweis von C. burnetii oder Coxiellose ist laut Verordnung über meldepflichtige Tierkrankheiten bei Wiederkäuern meldepflichtig (TKrMeldpflV 2015). Dabei sind die Leiter der Veterinäruntersuchungsämter, der Tiergesundheitsämter oder sonstiger öffentlicher oder privater Untersuchungsstellen sowie Tierärzte, die in Ausübung ihres Berufes meldepflichtige Krankheiten feststellen, verpflichtet das Auftreten der Krankheit oder des Erregers unverzüglich der nach Landesrecht zuständigen Behörde unter Angabe des Datums der Feststellung, der betroffenen Tierarten, des betroffenen Bestandes und des Kreises oder der kreisfreien Stadt zu melden. Eine Meldung von lediglich Antikörper-positiven Tieren ist demnach nicht gefordert, es sei denn diese werden in Zusammenhang mit Symptomen, die auf eine Coxiellose hindeuten, nachgewiesen. Die gemeldeten Fälle werden von den zuständigen Veterinärämtern in das Tierseuchen-Nachrichtensystem (TSN) eingetragen. Im Zeitraum von 2000 bis 2018 wurden weit mehr Fälle bei Rindern (8359 gemeldete Nachweise) als bei Schafen (1349 gemeldete Nachweise) und Ziegen (212 gemeldete Nachweise) gemeldet (Abb. 6). Dies könnte mehrere Gründe haben. Erstens führt möglicherweise trotz der gesetzlichen Vorgaben die zum Teil unterschiedliche Meldepraxis zu einer Verzerrung der Daten. Eine weitere Ursache ist vermutlich, dass in diesem Zeitraum weit mehr Rinder (4.100.863 Milchkühe ≥ 2 Jahre, DESTATIS 2018a) als Schafe (1.098.700 weibliche Schafe zur Zucht, DESTATIS 2018b) und Ziegen (88.451 weibliche Ziegen, statistische Erfassung alle drei Jahre seit 2010, DESTATIS 2016) in Deutschland gehalten wurden. Zum anderen ist das Bewusstsein von C. burnetii als möglicher Verursacher von Reproduktionsstörungen in der Rinderhaltung gestiegen (Lehner et al. 2017). Weiterhin ist bei der Interpretation der Meldedaten zu beachten, dass es in Deutschland kein aktives Überwachungssystem für Wiederkäuer gibt, mit dem die Prävalenz infizierter Tiere oder Herden repräsentativ überwacht werden könnte. Daher muss davon ausgegangen werden, dass die tatsächliche Anzahl betroffener Betriebe über der Meldezahl liegt. In Baden-Württemberg, Hessen, Niedersachsen, Nordrhein-Westfalen, Rheinland-Pfalz, Sachsen und Thüringen werden unter bestimmten Bedingungen die Kosten ganz oder zumindest teilweise für die C. burnetii-Impfung von der Tierseuchenkasse übernommen (Landwirtschaftskammer Nordrhein-Westfalen 2014, Lehner et al. 2017, Tierseuchenkasse Baden-Württemberg 2018, Tierseuchenkasse Hessen 2016, Tierseuchenkasse Rheinland-Pfalz 2017, Tierseuchenkasse Sachsen 2015, Tierseuchenkasse Thüringen 2019). Voraussetzung ist meist der molekularbiologische Nachweis des Erregers im tierischen Material. Außerdem wird in Bayern seit längerem zu Q-Fieber beim Rind geforscht (Boettcher et al. 2017, Böttcher et al. 2011b, 2013, 2019). Diese beiden Aktivitäten könnten in einzelnen Bundesländern zu höheren Meldezahlen beim Rind beigetragen haben. Bei kleinen Wiederkäuern liegen hauptsächlich Meldungen aus Süddeutschland vor (Abb. 6). Insgesamt befinden sich mehr Schaf- und Ziegenbetriebe in Süddeutschland als im Norden. Zusätzlich kam es in der Vergangenheit vermehrt zu humanen Kleinraumepidemien in Baden-Württemberg, Bayern, Hessen und Nordrhein-Westfalen, verursacht durch lammende Schafe. Deshalb ist es wahrscheinlich, dass das Bewusstsein für C. burnetii in diesen Regionen höher ist, wodurch gezielter auf diesen Erreger untersucht wird. Zudem ist die Zahl der weiblichen Ziegen von 2010 bis 2016 um 17 % auf 88.451 Tiere angestiegen (DESTATIS 2016). Gleichzeitig hat nach eigenen Beobachtungen eine Verschiebung von Kleinstbetrieben zu intensiven großen Milchziegenhaltungen mit mehreren hundert Tieren stattgefunden. Zusätzlich werden vermehrt kleinere Gruppen von Ziegen in großen Schafherden zur Landschaftspflege eingesetzt. Deshalb ist mit vermehrten Meldungen von C. burnetii bei dieser Tierart in den nächsten Jahren zu rechnen. Eine ähnliche strukturelle Änderung hatte in den 2000er Jahren in den Niederlanden stattgefunden. Retrospektiv betrachtet, hatte die höhere Tierzahl pro Betrieb jedoch nicht alleine das Risiko für eine C. burnetii-Infektion erhöht (Hogerwerf et al. 2013, Schimmer et al. 2011). Andere Faktoren wie beispielsweise die Dichte der Rinder haltenden Betriebe in der Region, die Durchführung der künstlichen Besamung bei Ziegen sowie der Zugang von Katzen und Hunden zum Ziegenstall haben ebenfalls eine Rolle gespielt (Hogerwerf et al. 2013, Schimmer et al. 2011).
In Ermangelung eines flächendeckenden und systematischen aktiven Überwachungssystems für C. burnetii muss bei der Frage nach der Verbreitung des Erregers auf aktive Studien in deutschen Schafherden zurückgegriffen werden. Hierzu wurden in den letzten 20 Jahren nur wenige durchgeführt. Informationen zu Ziegenherden oder Mischbetrieben (Schafe und Ziegen) sind besonders spärlich, obwohl in der Vergangenheit von ziegenassoziierten Q-Fieber-Fällen in Nord- und Süddeutschland berichtet wurde (Ganter et al. 2009, Sting et al. 2013). Die Datenerhebung bei Schafen beschränkt sich auf wenige Bundesländer. In Thüringen wurde eine Herdenprävalenz von 28 % (n = 39) mittels ELISA detektiert (Hilbert et al. 2012). In der gleichen Studie wurden auch Vaginaltupfer mittels PCR untersucht. Hierbei waren nur 5 % der Herden (n = 39) positiv. Zu einem späteren Zeitpunkt konnte bei 21,2 % (n = 99) der untersuchten Schafherden in Thüringen serologisch eine C. burnetii-Infektion nachgewiesen werden (Moog et al. 2016). In derselben Studie wurden auch 28 Ziegenherden auf Antikörperaktivität gegen C. burnetii getestet. Hierbei waren 14,3 % der Herden positiv. Insgesamt waren Schaf- und Ziegenherden mit einer Tierzahl > 399 häufiger serologisch positiv als kleinere Betriebe. Im Jahr 2004 wurden 95 niedersächsische Schafherden unterschiedlichster Größe ebenfalls serologisch auf Antiköper gegen C. burnetii untersucht (Runge et al. 2012). Hierbei konnte in 9,5 % der untersuchten Herden mindestens ein positives bzw. fraglich positives Tier gefunden werden. Außerdem wurde eine Einzeltierprävalenz von 2,7 % (n = 1714) für Niedersachsen ermittelt. Besonders auffällig war der hohe Anteil positiv getesteter Schafe pro Betrieb in drei Wanderschafherden. Sting et al. (2004) berichteten über einen Nachweis seropositiver Schafe von durchschnittlich 8,7 % (n = 3460) aus Baden-Württemberg. Für diese Untersuchung wurden Seren analysiert, welche ursprünglich 2001 für das Brucellose-Monitoring genommen wurden. In einer weiteren Untersuchung in neun Gemeinden Baden-Württembergs wurden 7,5 % der Schafe (n = 1036) positiv auf C. burnetii-Antikörper getestet (Brockmann 2014). Mittels phasenspezifischem ELISA wurden 2008 insgesamt 1187 Schafe aus 48 bayerischen Herden untersucht (Böttcher et al. 2011a). Dabei wiesen 9,8 % der analysierten Tiere Ph-I-Antikörper und 17,4 % der Schafe Ph-II-Antikörper auf. In den beiden darauffolgenden Jahren (2009: n = 997; 2010: n = 1032) lag die Nachweisrate sowohl für Ph-I-Antikörper (2009: 0,5 %; 2010: 0,6 %) als auch für Ph-II-Antikörper (2009: 0,8 %; 2010: 2,0 %) deutlich niedriger. Die vergleichsweise hohen Werte in 2008 lassen sich auf einen Q-Fieber-Ausbruch im Regierungsbezirk Unterfranken zurückführen. Aufgrund der unterschiedlichen Studiendesigns und der verwendeten Tests lassen sich die Ergebnisse der durchgeführten Studien nur eingeschränkt vergleichen.
Der direkte und indirekte Nachweis einer akuten Infektion mit C. burnetii beim Menschen ist gemäß Infektionsschutzgesetz (IfSG) namentlich meldepflichtig (IfSG 2019). Die Meldungen werden seit 2001 mit dem Überwachungsprogramm „SurvStat“ am Robert Koch-Institut (RKI) erfasst. Ende der neunziger Jahre kam es zu einem stetigen Anstieg von gemeldeten humanen Q-Fieber-Fällen, deren Anzahl sich seit den 2000er Jahren auf einem konstanten Niveau hält (Hellenbrand et al. 2001). Grundsätzlich besteht ein Unterschied in der Anzahl gemeldeter humaner Fälle zwischen Nord- und Süddeutschland. Teilweise lassen sich die Spitzen auf Kleinraumepidemien zurückführen (Abb. 7). Die drei größten Kleinraumepidemien wurden aus den Landkreisen Soest und Heidenheim an der Brenz sowie aus der Stadt Jena berichtet. So infizierten sich im Mai 2003 auf einem Bauernmarkt in der Nähe der Stadt Soest in Nordrhein-Westfalen zahlreiche der ca. 10.000 Besucher. 299 Besucher erkrankten an Q-Fieber (Porten et al. 2006). Aufgrund epidemiologischer Untersuchungen mittels ELISA konnte ein Texelschaf, das auf der Ausstellung lammte und lebende Zwillinge gebar, als Quelle der Infektion ausfindig gemacht werden. Dieses Mutterschaf stammte aus einer Herde mit einer Nachweisrate von 25 % serologisch positiver Schafe (n = 67). Im Juni 2005 waren in Jena eine kleine Gruppe von erstlammenden Merinoschafen, die in der Nähe einer Wohnsiedlung lammten, für die Infektion von 331 Personen verantwortlich (Gilsdorf et al. 2008). Auf einem Schafhoffest im Landkreis Heidenheim an der Brenz in Baden-Württemberg haben sich 228 Besucher im Juni 2014 mit C. burnetii infiziert (Fischer et al. 2016). Nur bei 20 % der untersuchten Patienten wurde eine asymptomatische Infektion diagnostiziert. Bei fünf Personen (2,2 %) bestand der Verdacht auf eine chronische Infektion.
Aufgrund der häufig nur subklinischen Verläufe ist von einer hohen Dunkelziffer bei der Erfassung von Q-Fieber-Infektionen beim Menschen auszugehen. Epidemiologische Auswertungen des niederländischen Ausbruchs legen den Verdacht nahe, dass nur etwa jede zehnte Infektion erkannt worden war (van der Hoek et al. 2012).
Die epidemiologische Situation von Q-Fieber-Infektionen für den Zeitraum 1947 bis 1999 wurde von Hellenbrand et al. (2001) evaluiert. Die Autoren zogen aus ihren Recherchen den Schluss, dass C. burnetii bei Rindern in vielen Gebieten Deutschlands seit den 1980er Jahren endemisch ist. Dagegen konzentriert sich das Vorkommen bei Schafen laut Hellenbrand et al. (2001) auf wenige Bundesländer im Süden, Osten und Westen Deutschlands. Seit 2000 gab es die meisten Nachweise sowohl bei kleinen Wiederkäuern als auch beim Menschen in Baden-Württemberg, Bayern, Hessen und Nordrhein-Westfalen. Thüringen hatte zwar 2005 eine humane Kleinraumepidemie und es wurden Nachweisraten von serologisch positiven Ziegen- und Schafherden zwischen 14,3 % und 28 % festgestellt (Hilbert et al. 2012, Moog et al. 2016), jedoch kam es mit Ausnahme von 2005 nur zu wenigen Meldungen bei Menschen und kleinen Wiederkäuern. Dies ist vermutlich auf die Bezuschussung des Impfstoffes für Schafe und Ziegen durch die thüringische Tierseuchenkasse und die damit vermehrte Impfung von Schaf- und Ziegenherden zurückzuführen (Moog et al. 2016, Tierseuchenkasse Thüringen 2019). Sicherlich spielen mehrere Faktoren eine Rolle für das Auftreten von Q-Fieber-Kleinraumepidemien beim Menschen. So konnten Frank und Höhle (2011) eine positive Korrelation zwischen der Schafdichte pro Hektar landwirtschaftlich genutzte Fläche und humanem Q-Fieber feststellen. Keinen Zusammenhang fanden die Autoren jedoch in Bezug auf Rinder und Ziegen. Von der räumlichen Korrelation zwischen Tierdichte und humanen Q-Fieber-Fällen kann zwar noch keine Kausalitätsvermutung abgeleitet werden, jedoch kann dieses Ergebnis als Grundlage für weiterführende Studien dienen. Weiterhin ist auffällig, dass die drei großen Epidemien seit 2000 ihren Anfang im Frühsommer, einer warmen und trockenen Jahreszeit, hatten. In dieser Jahreszeit lammen besonders asaisonale Rassen wie Merinoschafe im Freien. Die meisten asaisonalen Rassen werden bevorzugt in Süddeutschland gehalten. Im Gegensatz dazu ist die größte Schafsdichte pro landwirtschaftlich genutzter Fläche in Schleswig-Holstein zu finden. Jedoch gibt es sowohl in der Human- als auch in der Veterinärmedizin nur wenige Coxiellen-Infektionsnachweise in diesem Bundesland. Dies könnte an der saisonalen Lammung der Schafe im Stall in den Monaten Februar und März liegen, einer besonders feuchten und noch kalten Jahreszeit. Zusammenfassend könnte die asaisonale Lammung in der warmen und trockenen Jahreszeit sowie eine hohe Schafdichte zu vermehrten Q-Fieber-Infektionen führen. Hellenbrand et al. (2001) sahen auch das Vorhandensein von D. marginatus in Süddeutschland als einen Risikofaktor an. Jedoch kommt diese Zeckenart nicht in Nordrhein-Westfalen vor (Rubel et al. 2014, Walter et al. 2016). Treffen die genannten Risikofaktoren auf eine humane Population (Wohngebiet, Marktbesucher, etc.), kann dies zu Erkrankungsfällen bzw. Kleinraumepidemien führen. Diese Annahmen gilt es zukünftig zu überprüfen. Außerdem dürfte mit dem Anstieg der professionellen Ziegenhaltung in Deutschland diese Tierart zunehmend eine Rolle bei der Q-Fieber-Epidemiologie spielen. Sowohl in Bulgarien als auch in den Niederlanden konnte ein Zusammenhang zwischen der Zunahme an Ziegen und steigenden humanen Q-Fieber-Erkrankungen festgestellt werden (Mori und Roest 2018).
Diagnostik
Serologie
Für den serologischen Nachweis einer C. burnetii-Infektion fehlt in der Veterinärmedizin ein Goldstandard (van den Brom et al. 2015c). Zudem ist die veterinärmedizinische Q-Fieber-Serologie noch weit von einer Standardisierung entfernt (Rousset et al. 2007). In der Routinediagnostik wurde lange Zeit die Komplementbindungsreaktion (KBR) als Methode der Wahl zum Nachweis von C. burnetii-Antikörpern eingesetzt. Diese Methode verwendet grundsätzlich eine Mischung aus Ph-I- und Ph-II-Antigenen vom Nine-Mile-Stamm (OIE 2018). In der KBR gelten bei Wiederkäuern Titer zwischen 1:10 und 1:40 als latente Infektionen. Titer von 1:80 und darüber werden als akute Phase der Infektion eingestuft (OIE 2018). Mittlerweile wurde die KBR als Standardmethode vom ELISA abgelöst (OIE 2018), da in Vergleichsuntersuchungen die KBR eine geringere Sensitivität aufweist als die verwendeten ELISAs (Kittelberger et al. 2009, Natale et al. 2012, Rousset et al. 2007, Sting et al. 2004). Schmeer (1985) begründet dies mit der partiellen Hemmung des komplementbindenden IgG1 durch IgG2 und IgM, die sich in der KBR als Titerreduktion und unvollständige Hemmung der Hämolyse äußert. Infolgedessen können besonders in niedrigen Titerbereichen mit der KBR falsch negative Ergebnisse zu Stande kommen (Schmeer 1985). Allerdings bindet in der KBR auch IgM ein Komplement. Deshalb kann mittels KBR, im Gegensatz zu den IgG-basierten ELISAs, die frühe humorale Immunantwort auch detektiert werden (Kittelberger et al. 2009). Kommerziell erhältliche ELISAs basieren auf einer Mischung von Ph-I- und Ph-II-Antigenen und detektieren nur IgG-Antikörper. Die aktuell in Deutschland entsprechend dem Tiergesundheitsgesetz (TierGesG 2019) zugelassenen ELISAs lassen keine sichere Aussage über das Infektionsgeschehen innerhalb einer Herde zu. Sie sind aber aufgrund ihrer guten Automatisierbarkeit zum Untersuchen großer Probenzahlen geeignet (OIE 2018). Validiert wurden die ELISAs meist auf der Grundlage der KBR, ohne klar definierte Negativ- und Positivkontrollen. Die ELISAs lieferten mehr positive Reagenten als die KBR, dies wurde als eine höhere Sensitivität interpretiert. Ein positives Ergebnis mit der KBR oder dem ELISA gibt jedoch lediglich Aufschluss darüber, ob das Tier Kontakt mit C. burnetii hatte und es zu einer humoralen Immunantwort kam. Beide Tests lassen keine Aussage über die Erregerausscheidung zu. Serologisch negativ getestete Tiere können den Erreger ausscheiden (de Cremoux et al. 2012, Joulié et al. 2017, Natale et al. 2012, Rodolakis et al. 2007). Bei einem Infektionsversuch von Arricau-Bouvery et al. (2003) kam es zu ersten Aborten und somit zur Erregerausscheidung 12 Tage bzw. 25 Tage nach subkutaner Infektion mit C. burnetii. Erst 42 Tage nach der experimentellen Infektion wurden die ersten Ziegen mittels ELISA serologisch positiv getestet. Deshalb sind die serologischen Tests alleine nicht dazu geeignet die Infektionsfreiheit von Einzeltieren und Herden zu bestätigen (Angelakis und Raoult 2010, OIE 2018). Jedoch können sie über das Vorhandensein von Antikörperaktivitäten auf Bestandsebene Aufschluss geben. Um Einzeltiere bzw. Herden kleiner Wiederkäuer als frei von C. burnetii zu testen, liefert aktuell nur die PCR-Untersuchung, bevorzugt von Pueperaltupfern, zuverlässige Ergebnisse (OIE 2018).
Um einen tieferen Einblick in das Infektionsgeschehen in Herden zu erhalten, kann ein phasenspezifischer Nachweis sowohl mit dem indirekten Immunfluoreszenz-Antikörpertest (IFAT) als auch mit phasenspezifischen ELISAs durchgeführt werden. Beide Testverfahren sind derzeit in Deutschland nach dem Tiergesundheitsgesetz (TierGesG 2019) für die Diagnostik nicht zugelassen und werden bislang für wissenschaftliche Zwecke eingesetzt. Bisher wurden mit diesen Tests vor allem Serumproben von Rindern und Ziegen untersucht (Böttcher et al. 2011b, Muleme et al. 2016, Roest et al. 2013, Sting et al. 2013). In einem Infektionsversuch mit Ziegen konnte gezeigt werden, dass sowohl IgM Ph-II- als auch IgG Ph-II-Antikörper im phasenspezifischen ELISA zeitgleich ab der zweiten Woche nach intranasaler Inokulation von C. burnetii ansteigen (Roest et al. 2013). Zum gleichen Zeitpunkt kam es auch zu einem Anstieg von IgM Ph-I-Antikörpern, jedoch in einem geringeren Umfang. IgG Ph I stiegen ab der 5. Woche nach Infektion langsam aber stetig an. Daraus postulierten Roest et al. (2013), dass ein Anstieg von IgM Ph II in Abwesenheit von IgG Ph-I-Antikörpern für eine akute Infektion bei Ziegen sprechen. Ab der 10. Woche post infectionem waren sowohl IgG Ph-II- als auch IgG Ph-I-Antikörper auf demselben hohen Niveau. Zu einem ähnlichen Ergebnis kamen auch Sting et al. (2013) bei der Untersuchung eines Q-Fieber-Ausbruchs bei Milchziegen. Sie definierten eine akute Infektion besonders mit dem Vorhandensein von IgG Ph-II-Antikörpern und nur schwachen Signalen von IgG Ph-I-Antikörpern. IgG Ph-I-Titer sind bei Ziegen über zwei Jahre nachweisbar, während IgG Ph-II-Titer zu diesem Zeitpunkt meistens nicht mehr nachweisbar sind (Hatchette et al. 2003). Ausführliche Langzeitstudien zum phasenspezifischen Verlauf von IgM und IgG fehlen sowohl für Ziegen als auch bei Schafen. Obwohl umfangreiche Validierungen der phasenspezifischen Tests noch ausstehen, scheint die phasenspezifische Serologie ein wertvolles Diagnostikinstrument zu sein, um die Infektionsdynamik in Herden zu analysieren und Neuinfektionen zu detektieren (Muleme et al. 2017). Jedoch sind weitere Studien über einen längeren Zeitraum mit gezielten Infektionen für die einzelnen Wiederkäuerspezies notwendig, um verbindlichere Aussagen treffen zu können. Zudem fehlen klare Falldefinitionen bezüglich des Infektionsstatus (frei, akut bzw. chronisch infiziert) in der Veterinärmedizin. Eine Unterscheidung zwischen einem geimpften oder natürlich infizierten Tier ist mit keiner serologischen Methode möglich (EFSA 2010).
Neben Serumproben können auch Einzelmilchproben oder Tankmilchproben durch ELISA untersucht werden. Mittels Tankmilchproben konnten bei einer Inner-Herdenprävalenz von mindestens 10–15 % eine Infektion mit C. burnetii in Schaf- und Ziegenherden mit dem ELISA nachgewiesen werden (Ruiz-Fons et al. 2011, van den Brom et al. 2012). Bei der Verwendung von Tankmilchproben gelang der Nachweis mittels ELISA häufiger als mit der PCR (Muleme et al. 2017, van den Brom et al. 2012). Die zellulär vermittelte Immunität gegen C. burnetii lässt sich durch Interferon-γ-(IFN-γ)-recall- oder -release-Tests ermitteln. Während einer Infektion werden Erreger durch Antigen-präsentierende Zellen (APCs) aufgenommen und über die Präsentation der Erregerantigene auf der Zelloberfläche zusammen mit kostimulierenden Signalen T-Lymphozyten aktiviert. Aktivierte TH1-Zellen schütten dann bei erneutem Erregerkontakt IFN-γ aus. Dies macht man sich in einem IFN-γ-recall-Test zunutze. Wurden TH1-Zellen des zu untersuchenden Tieres schon einmal während einer Infektion mit C. burnetii-spezifischen Antigenen konfrontiert, kommt es zu einer starken Produktion von IFN-γ. Mit diesem Test kann eine zelluläre Immunreaktion nachgewiesen werden. Dieses Testverfahren wurde in der Veterinärmedizin zum Nachweis einer C. burnetii-Infektion bisher noch wenig eingesetzt (Boettcher et al. 2017, Janowetz et al. 2018, Roest et al. 2013). Für die humanmedizinische Diagnostik wurde der Nachweis der T-Zellimmunität gegenüber Coxiellen ebenfalls etabliert (Limonard et al. 2012). Er kann als ergänzende Methode zur Diagnostik des chronischen Q-Fiebers und dessen Therapiekontrolle verwendet werden.
Molekularbiologie
Zum direkten Nachweis von C. burnetii wird heutzutage zumeist eine PCR verwendet. Aktuell gibt es in Deutschland vier zugelassene real-time PCR-Testkits für die Untersuchung von veterinärmedizinischen Proben, mit denen ein repetitiv im Genom vorkommendes Transposase-Element (IS1111) von C. burnetii amplifiziert wird. Diese PCRs können beispielsweise zum Nachweis von C. burnetii-spezifischer DNS aus Plazentaproben, Tupfern von Vaginalschleimhaut sowie Milchproben von Hauswiederkäuern verwendet werden. Abhängig vom Coxiellen-Isolat kann dieses repetitive Element zwischen 7- und 110-mal pro Genom je Bakterienzelle vorkommen. Bei den meisten Isolaten kommt die Insertionssequenz jedoch zwischen 10- und 30-mal vor (Klee et al. 2006). Dadurch ist eine PCR-Diagnostik, die auf dem Nachweis der IS1111-Region beruht, äußerst sensitiv. Eine exakte Quantifizierung der Erregermenge ist aufgrund der variablen Anzahl der Insertionselemente bei unterschiedlichen C. burnetii-Stämmen nicht möglich, allerdings im Rahmen von Verlaufsuntersuchungen zur Erregermengenausscheidung von untergeordneter Bedeutung. Exakte Erregerquantifizierungen sind mithilfe von PCRs, deren Zielsequenzen single-copy-Gene wie com1 verwenden, möglich (Kersh et al. 2010).
Die sogenannte Multiple Loci Variable Number of Tandem Repeats (VNTR) Analysis (MLVA) ermöglicht die Typisierung von gesamten Genomsequenzen mit hoher diskriminierender Kraft (Arricau-Bouvery et al. 2006). Für Deutschland konnten dadurch zwei Haupt-Cluster innerhalb der Wiederkäuerspezies identifiziert werden (Frangoulidis et al. 2014). Cluster A wurde besonders aus Isolaten von kleinen Wiederkäuern detektiert, wobei das Subcluster A1 eher ziegenassoziiert ist und A2 vor allem bei Schafen nachgewiesen wurde. Bei Isolaten aus Rinderproben wurde primär das Cluster C nachgewiesen. Zusätzlich gab es auch unterschiedliche geographische Verteilungen dieser Cluster. Somit dürfte es innerhalb der Wiederkäuerspezies unterschiedliche Isolate-Gruppen in verschiedenen Regionen geben. Über Wirtsspezifität, Pathogenität und Virulenz des jeweiligen Clusters lassen sich jedoch bislang noch keine Aussagen treffen.
Neben der VNTR/MLVA-Methode wird auch noch das SNP-basierte Multispacer-Sequence-Typing (MST)-Verfahren regelmäßig verwendet (Glazunova et al. 2005). Allerdings erreicht dieses Verfahren nur eine sehr grobe Auflösung in Bezug auf regionale Verteilungen (Massung et al. 2012).
Für das Monitoring von Milchziegen- und Milchschafbeständen mittels Tankmilchproben haben sich neben der Anwendung von ELISAs vor allem auch die PCR-Methoden bewährt (Ruiz-Fons et al. 2011, Sidi-Boumedine et al. 2010, van den Brom et al. 2012, 2015b.). Mit der PCR lassen sich Intra-Herden-Seroprävalenzen ab 15 % detektieren (van den Brom et al. 2012). Die Probenentnahme sollte jedoch frühestens zwei Wochen nach einer Impfung mit C. burnetii-Antigenen stattfinden, da Impfstoff assoziierte C. burnetii-DNS bis zu 9 Tage nach Impfung über die Milch ausgeschieden wird (Hermans et al. 2011). Eine zuverlässige und aktive Surveillance fehlt weltweit, insbesondere für nicht milchproduzierende Schaf- und Ziegenbestände. Deshalb könnte ein möglicher Ansatz die Verwendung von Umweltproben sein. Infizierte Tiere scheiden während eines Abortes bzw. einer Normalgeburt C. burnetii massiv aus und kontaminieren somit ihre Stallungen. Staub von Fensterbänken und Stalleinrichtungen können zum Nachweis von C. burnetii in der Tierhaltung verwendet werden (Carrié et al. 2019, Joulié et al. 2015). In einer Studie von Carrié et al. (2019) konnten, besonders in Herden kleiner Wiederkäuer, Coxiellen mittels PCR nachgewiesen werden, teilweise mit bis zu 108 Genomäquivalenten. Die Wahrscheinlichkeit, dass der Erreger im Stallstaub nachgewiesen wurde, stieg mit der Zunahme an Muttertieren. Auch Stallluftproben (Astobiza et al. 2011a, Joulié et al. 2015) und Dungproben (Avbersek et al. 2019) wurden zum Nachweis von C. burnetii in Tierhaltungen verwendet. Bei einem positiven Nachweis mit der PCR kann jedoch keine Aussage getroffen werden, ob es sich um ein akutes Q-Fieber-Geschehen handelt oder um eine Erregerausscheidung in der Vergangenheit. Umweltproben können über einen längeren Zeitraum positiv bleiben, auch wenn keine Erregerausscheidung mehr stattfindet (Astobiza et al. 2011b). Weiterhin ist es in der Routinediagnostik nicht möglich, eine Aussage über die Lebensfähigkeit der Coxiellen zu treffen. Álvarez-Alonso et al. (2018) konnten mittels aufwendiger Inokulation in Mäusen vermehrungsfähige Coxiellen zwei Monate nach der letzten Lammung in Staubproben nicht mehr nachweisen. Auch van den Brom et al. (2015a) gelang es nicht, Coxiellen aus Ziegenmist, der bereits drei Monate gelagert wurde, zu kultivieren. Deshalb gehen die Autoren davon aus, dass sich in ihren Proben keine lebensfähigen Coxiellen mehr befanden. Diese Ergebnisse gilt es in Zukunft in weiteren Studien zu überprüfen und die Tauglichkeit von Umweltproben als Probenmatrix zu evaluieren. Bei wiederholtem negativem Ergebnis aus Staubproben kann davon ausgegangen werden, dass die Tiere keine Coxiellen ausscheiden. Zusätzlich könnten Oberflächenproben als Marker nach der Reinigung und Desinfektion von Stallungen verwendet werden, um den Erfolg dieser Maßnahmen zu kontrollieren.
Um die Ausscheidung in Herden kleiner Wiederkäuer zu überwachen, können auch Vaginaltupfer verwendet werden. Besonders in geimpften Herden kann damit eine Aussage über die Erregerausscheidung und somit des Impferfolgs getroffen werden. Diese sollten nicht später als 8 Tage nach einem Abort bzw. einer Geburt genommen werden (OIE 2018). Die Anzahl und der Zeitpunkt der Probenentnahme variieren je nach Autor stark. Beispielsweise wird unabhängig von der Herdengröße eine Beprobung von 10 Muttertieren mit Vaginaltupfern empfohlen, die innerhalb der letzten drei Monate gelammt haben (CVUA Stuttgart 2018). Im Gegensatz dazu empfehlen de Cremoux et al. (2018), ebenfalls unabhängig von der Herdengröße, die Beprobung von nur zwei bis sechs Tieren mittels Vaginaltupfern die innerhalb von einer Woche abortiert haben.
Van Leuken et al. (2016) empfehlen zur Überwachung eines Q-Fieber-Geschehens eine Kombination aus Luftproben innerhalb und außerhalb des Stalls mit Vaginaltupfern und Tankmilchproben. Die Vielzahl an Empfehlungen zur Überwachung macht deutlich, dass die Standardisierung einer aktiven Surveillance unbedingt notwendig ist. Bei der Verwendung molekularbiologischer Nachweise hat bereits eine Erregerausscheidung stattgefunden. Demzufolge fehlt eine aktive Surveillance, die positive Herden frühzeitig detektiert, bevor C. burnetii ausgeschieden wird.
Maßnahmen
Sofortmaßnahmen
Liegt ein akutes Infektionsgeschehen in Ziegen- und Schafbeständen vor, dann sollten verschiedene Maßnahmen getroffen werden, um die Gefahr einer Infektion der Bevölkerung weitestgehend zu minimieren. Unter anderem lassen sich solche Schritte in den Empfehlungen für Hygienemaßnahmen bei der Haltung von Wiederkäuern (BMEL 2014) und im Leitfaden zum Q-Fieber in Baden-Württemberg (CVUA Stuttgart 2018) sowie in Lehrbüchern (Selbitz und Ganter 2018) finden. Lammungen und Schur sollten nur in geschlossenen Räumen stattfinden und Nachgeburten sowie Aborte müssen in geschlossenen Behältnissen bis zur Abholung durch Verarbeitungsbetriebe Tierischer Nebenprodukte aufbewahrt werden. Betriebsfremde Personen dürfen keinen Zugang zu den Tieren haben. Rohmilch und deren Produkte sollten weder verkauft noch verzehrt werden. Die Milch ist zu pasteurisieren. Weiterhin sind Arbeitskleidung sowie Arbeitsmaterialien zu reinigen und zu desinfizieren. Die häufig geforderte Verwendung von Branntkalk (ca. 10 kg/m2, Kalkhydrat, Ca(OH)2, BMEL 2007) zur Erstellung einer Festmistpackung wird von den Autoren im Falle der C. burnetii-Bekämpfung aufgrund der Gefahr der Selbstentzündung nicht geteilt. In einer niederländischen Studie konnte 97 Tage nach Lagerung von großen Ziegenfestmistpackungen (mindestens 10 Meter lang, 4,5 Meter breit und 3,5 Meter hoch) unter der Folie zwar noch C. burnetii-DNS nachgewiesen werden, die Anzucht des Erregers gelang jedoch nicht mehr (van den Brom et al. 2015a). Deshalb sollte der Mist von infizierten Beständen nicht sofort ausgebracht, sondern für mindestens drei Monate unter Folie sowie abseits von Wohngebieten gelagert werden (Plummer et al. 2018). Anschließend ist der Mist bei windstillem Wetter auf Ackerflächen auszubringen und sofort in den Boden einzuarbeiten. Weiterhin wird gefordert Dauerausscheider in einem Bestand beispielsweise mittels Scheidentupfer zu identifizieren, um diese anschließend zu töten und unschädlich zu beseitigen (BMEL 2014). Einzelne mit C. burnetii infizierte Ziegen scheiden den Erreger auch nach einer Impfung intermittierend über die Milch aus (van den Brom et al. 2013). Diese Ausscheider sollten über Einzelmilchproben erkannt und geschlachtet werden. Bei diesen Ziegen wurde C. burnetii-DNS nur im Euter nachgewiesen (van den Brom et al. 2013). Deshalb sollte bei der Schlachtung das Euter unversehrt abgenommen und verworfen werden. Dauerausscheider wurden bei Schafen bisher nicht identifiziert (Astobiza et al. 2011b). Eine Chronifizierung der C. burnetii-Infektion wurde auch beim Schaf bisher noch nicht nachgewiesen. Im Rahmen der Bekämpfungsmaßnahmen ist eine enge Kooperation und Koordination zwischen den Gesundheitsämtern, den Veterinärämtern, den Hausärzten und Tierärzten sowie den Tiergesundheitsdiensten und insbesondere den betroffenen Tierhaltern anzustreben (Selbitz und Ganter 2018).
Antibiotikabehandlung
Der Einsatz von Oxytetracyclin zur Behandlung von Coxiellosen führte nicht zu einer Reduzierung der Ausscheidung bei kleinen Wiederkäuern (Astobiza et al. 2013, de Cremoux et al. 2012). Allerdings ist der Einsatz von Oxytetracyclin im Ausbruchsgeschehen mit einer gleichzeitigen Infektion mit Chl. abortus gerechtfertigt (Eibach et al. 2013). Im Gegensatz zu kleinen Wiederkäuern schieden Rinder nach der Behandlung mit Oxytetracyclin weniger Coxiellen aus (Taurel et al. 2012). Aufgrund des sehr niedrigen pH-Werts in den Phagolysosomen der Wirtszelle, in denen sich C. burnetii vermehrt, kommt es zur Inaktivierung von Tetracyclinen durch Epimerisierung am C4-Atom (Rogalski 1985). Demgegenüber gilt Doxycyclin in der Humanmedizin als Mittel der Wahl bei der Behandlung des Q-Fiebers. Doxycyclin weist nicht nur höhere in-vitro-Aktivität und bessere pharmakokinetische Parameter auf (Rogalski 1985), sondern ist deutlich resistenter gegenüber niedrigen pH-Werten. Einen Therapieerfolg verhindern können Doxycyclin-resistente Stämme von C. burnetii, die in der Literatur beschrieben wurden (Rolain et al. 2005). Mechanismen, die für diese Resistenzen verantwortlich sein könnten, sind bislang nicht bekannt (Rouli et al. 2012). Über die Wirksamkeit anderer in der Tiermedizin verwendeter Antibiotika liegen aktuell keine Erkenntnisse vor.
Impfung
Seit 2010 gibt es in Deutschland einen für Rinder und Ziegen zugelassenen Impfstoff (Coxevac®, Ceva Santé Animale, Libourne, Frankreich). Dabei handelt es sich um einen inaktivierten Ph-I-Impfstoff. Dieser besitzt eine bessere Wirkung als Ph-II-Impfstoffe (Arricau-Bouvery et al. 2005). Der Mechanismus der protektiven Immunität von Ph-I-Impfstoffen konnte noch nicht vollständig aufgeklärt werden (Achard und Rodolakis 2017). Die Grundimmunisierung im Abstand von drei Wochen sollte vier Wochen vor der ersten Bedeckung abgeschlossen sein. Auf diese Weise wird die Ausscheidung von C. burnetii bei naiv geimpften Ziegen stark gesenkt, aber nicht völlig verhindert (Arricau-Bouvery et al. 2005). In bereits infizierten Ziegenherden wird mit der Impfung die Ausscheidung von C. burnetii langfristig verringert. Durch die Impfung von Jungtieren vor der ersten Bedeckung wird die Erregerausscheidung in positiven Betrieben besonders stark reduziert (Achard und Rodolakis 2017, de Cremoux et al. 2012, Rousset et al. 2009b). Dies könnte vermutlich daran liegen, dass C. burnetii für eine erfolgreiche Etablierung der Infektion Trophoblasten benötigt (Ben Amara et al. 2010, Roest et al. 2012, Sánchez et al. 2006). Eine Impfung von Jungziegen vor der ersten Bedeckung scheint deshalb die Etablierung von C. burnetii zu erschweren. Zur Aufrechterhaltung der Immunität müssen die Ziegen jährlich geboostert werden. Bei Ziegen kann eine Impfung zu einer zeitweise geringen bis moderaten Schwellung der Haut an der Injektionsstelle sowie kurzfristig zu Fieber (> 40,5 ºC) und Rückgang der Milchleistung führen (EMA 2014, Vellema et al. 2010). Die subkutane Schwellung ist bei bereits mit C. burnetii infizierten Ziegen massiver als bei naiven Tieren (Vellema et al. 2010). Der Impfstoff hat keine Zulassung für die Anwendung bei tragenden Tieren, dennoch wurde dieser in der Vergangenheit bei trächtigen Schafen (Eibach et al. 2013, Joulié et al. 2017) und Ziegen (persönliche Erfahrungen der Autoren Ganter und Bauer) eingesetzt. Teilweise konnte damit eine Reduzierung der Ausscheidung bei Schafen erreicht werden (Eibach et al. 2013). Im Gegensatz dazu konnten Astobiza et al. (2013, 2011a) keinen signifikanten Unterschied zwischen geimpften und ungeimpften Schafen bezüglich der Ausscheidung von C. burnetii über das Vaginalsekret feststellen. In ihrer ersten Studie wurden zwei C. burnetii positive Milchschafherden sechs bzw. drei Wochen vor der künstlichen Besamung mit einem Ph-I-Impfstoff geimpft. Ein Viertel der Mutterschafe blieb jeweils als Kontrollgruppe ungeimpft. Innerhalb von vier Wochen nach der Lammung wurden Vaginaltupfer genommen und mittels PCR untersucht. Im Vergleich zum Vorjahr ging insgesamt in beiden Herden unabhängig vom Impfstatus der Tiere die Erregerausscheidung signifikant zurück (Herde 1: von 6,5 log auf 3,4 log C. burnetii pro Tupfer, Herde 2: von 7,3 log auf 2,9 log C. burnetii pro Tupfer). Ein signifikanter Unterschied zwischen geimpfter und ungeimpfter Teilherde bestand jedoch nicht (Astobiza et al. 2011a). Dieses Ergebnis wurde in einer ähnlichen Studie der Autoren nochmals bestätigt (Astobiza et al. 2013). Trotz der fehlenden Zulassung des Impfstoffes für Schafe kann dieser jedoch speziesübergreifend angewendet werden (StIKoVet 2018). Eine Produktinformation des Herstellers Ceva aus dem Jahre 2008 empfiehlt eine Impfdosis von Coxevac® beim Schaf von 1 ml. Aktuelle Daten sind vom Hersteller dazu nicht erhältlich. In der Praxis wird seit Jahren sowohl national (Eibach et al. 2013, Hamann et al. 2009) als auch international (persönliche Kommunikationen Renée De Cremoux, Frankreich, Ana García-Peréz, Spanien und René van den Brom, Niederlande) die Impfdosis von 1 ml beim Schaf erfolgreich angewendet. Es handelt sich dabei jedoch nur um empirische Erfahrungswerte, die wissenschaftlich noch nicht nachgewiesen sind. Zusammenfassend ist es bislang nur mit der Impfung möglich, C. burnetii-Infektionen zu verhindern bzw. zu kontrollieren (Bontje et al. 2016). Deshalb sollte die Impfung als wichtiges Kontrollinstrument zum Einsatz kommen, um Herden vor C. burnetii zu schützen und humane Infektionen zu verhindern (Bontje et al. 2016, EFSA 2010).
Desinfektion
Aufgrund der hohen Umweltresistenz von C. burnetii sind viele Desinfektionsmittel nicht im ausreichenden Umfang wirksam. Besonders die unterschiedliche Tenazität der morphologischen Zellformen von C. burnetii, mit den vegetativen LCV und den Sporen-ähnlichen SCV, erschwert die Auswahl von geeigneten Desinfektionsmaßnahmen.
Generell fehlen genaue Informationen und Empfehlungen bezüglich Auswahl und Anwendung von Desinfektionsmitteln, um C. burnetii positive Tierhaltungsbereiche zu dekontaminieren (Frentzel et al. 2013, Rodolakis 2009). Sehr allgemein werden bei bakteriellen Tierseuchenerregern entsprechende Desinfektionsmittel aus der Liste der Deutschen Veterinärmedizinischen Gesellschaft e. V. (Spalte 4a, „spezielle Desinfektion“, DVG 2019), die in der Gebrauchskonzentration innerhalb von 2 Stunden als wirksam eingetragen sind, empfohlen (BMEL 2007). Konkrete Ausführungen über den Einsatz von Desinfektionsmitteln sind vielfältig. Das RKI (2012) empfiehlt für die Desinfektion von Stallungen 10–20%ige Chlorkalklösungen, eine 1%ige Lysol-Lösung oder 5%ige Wasserstoffsuperoxid-Lösungen. Bei der Verwendung einer 2%igen Natronlauge konnte auf Sand-, Lehm- und Lössboden eine Reduzierung der Keimlast um mindestens den Faktor 104 erreicht werden (Dörner 2011). Der Einsatz einer 1%igen Peroxygenverbindung führte nach 30 Minuten zu einer Reduzierung der Infektiosität von über 90 % (Priestley 2007). Nach intensiver Reinigung und anschließender Desinfektion mit 0,5%iger Peressigsäure konnte in einem Schafstall die Erregerlast von C. burnetii deutlich reduziert werden (Moog et al. 2018). Weiterhin gilt zu beachten, dass viele Stallungen von kleinen Wiederkäuern aus Naturmaterialen (z. B. Holzwände und -gatter sowie Lehmböden) bestehen. Diese lassen sich nicht ausreichend reinigen und desinfizieren. Deshalb kann nur eine Erregerreduzierung und keine Erregerfreiheit erreicht werden. Die Überprüfung von potenziell wirksamen Desinfektionsmitteln ist dringend erforderlich, um zukünftig evidenzbasierte und praxistaugliche Empfehlungen zur Desinfektion von C. burnetii kontaminierten Tierhaltungen geben zu können. Auch bei der Reinigung und Desinfektion von Stallungen sollten entsprechende Schutzmaßnahmen getroffen werden, um eine Infektion der durchführenden Personen und der Umgebung zu verhindern. Besonders sollte bei dieser Tätigkeit die Entwicklung von Aerosolen und Stäuben verhindert werden.
Für die Desinfektion von Kleidung werden Hitzebehandlungen (90 °C für 10–15 min) oder ein durch das RKI gelistetes Desinfektionswaschverfahren (RKI 2017) empfohlen.
Zoonose
Die Inkubationszeit nach aerogener Infektion beträgt beim Menschen durchschnittlich 18 Tage, reicht jedoch von 7 bis 32 Tagen (Todkill et al. 2018). Eine neuere Modellberechnung von Brooke et al. (2013) hat die humane Infektionsdosis (ID50), bei der 50 % der Exponierten sich durch kontaminierte Aerosole infizieren, auf 1,18 Bakterien (0,76–38,7; 95%-Konfidenzintervall) geschätzt. Dies verdeutlicht die hohe Kontagiosität von C. burnetii, besonders für den Menschen. Die Infektion verläuft bei ca. 60 % der Fälle symptomlos. Jedoch zeigen 40 % der infizierten Personen grippeähnliche Symptome wie hohes Fieber, Kopfschmerzen und respiratorische Erkrankungen (Maurin und Raoult 1999). Infizierte Personen entwickeln in bis zu 5 % der Fälle eine Chronifizierung (Wegdam-Blans et al. 2012), die sich meist als Endokarditis manifestiert (Botelho-Nevers et al. 2007, Million et al. 2010, Raoult et al. 2000). Weiterhin besteht ein nicht unerhebliches Risiko, nach einer akuten Infektion mit C. burnetii am chronischen Müdigkeitssyndrom (Q-Fieber-Fatigue-Syndrome, QFS) zu erkranken. Im Durchschnitt leiden 20 % der Patienten mit akuter Q-Fieber-Erkrankung anschließend an chronischer Müdigkeit, wobei dieser Krankheitskomplex bis zu 20 Jahre anhalten kann (Morroy et al. 2016). Die Behandlung einer akuten Q-Fieber-Erkrankung erfolgt primär für zwei Wochen mit Doxycyclin. Bei chronischen Erkrankungen wird Doxycyclin mit Hydroxychloroquin kombiniert, muss jedoch mindestens 18 Monate lang eingenommen werden. Für die Prophylaxe steht in Europa kein Impfstoff zur Verfügung. Kann eine Exposition mit C. burnetii nicht ausgeschlossen werden, so wird von dem U. S. Army Medical Research Institute of Infectious Diseases (USAMRIID) eine Postexpositionsprophylaxe empfohlen. Mit der Einnahme von Doxycyclin (100 mg per os, alle 12 Stunden) über 5 bis 7 Tage sollte etwa am 8. bis 12. Tag nach Exposition begonnen werden (USAMRIID 2014).
Ausblick
Trotz der Vielzahl an Q-Fieber-Ausbrüchen in den letzten 20 Jahren in Deutschland mit vielen humanen Fällen sind keine umfassenden Daten zur Epidemiologie und Prävention bei Mensch und Reservoir-Tieren vorhanden. Im Rahmen der Zoonose-Initiative der Bundesregierung sollen viele offene Fragen durch den Verbund „Q-GAPS: Q fever – German Interdisciplinary Program for Research“ bearbeitet werden. Das Ziel von Q-GAPS ist es, diese ungelösten Fragen zur Epidemiologie, Pathogenese, Überwachung und Kontrolle von C. burnetii im Sinne des One-Health-Gedankens zu adressieren. Die Ergebnisse werden neue Einblicke in die C. burnetii-Infektion bei kleinen Wiederkäuern geben. Ebenfalls wird die Rolle von Zecken bei der Übertragung von C. burnetii analysiert. Ein weiterer wichtiger Aspekt ist die Untersuchung des Vorkommens des Fatigue-Syndroms nach einer Q-Fieber-Infektion (QFS). Zudem sollen Desinfektionsmittel nach den Richtlinien der Deutschen Veterinärmedizinischen Gesellschaft und
des Verbundes für angewandte Hygiene geprüft und Empfehlungen erstellt werden. Weiterhin wird die Antigenpräsentation während der C. burnetii-Infektion erforscht,
um mögliche Impfstrategien zu entwickeln. Durch die Sequenzierung diverser Isolate und der Korrelation der Genome mit ihrer Virulenz sollen Gene identifiziert werden, die mit unterschiedlichen Wirtsspezifitäten und/oder dem pathogenen Potenzial assoziiert sein können. Durch die Analyse der Funktionsweise dieser Gene können mögliche Virulenz-Marker von C. burnetii identifiziert werden. Alle Ergebnisse werden in eine bereits bestehende, öffentlich zugängliche interaktive Datenbank einfließen (www.q-gaps.de) und in eine Q-Fieber-Leitlinie integriert, um den öffentlichen Gesundheitsdienst bei der Erkennung, Überwachung und Bekämpfung von C. burnetii-Infektionen zu unterstützen.
Conflict of interest
Die Autoren sind Mitglied im Zoonosekonsortium Q-GAPS (www.q-gaps.de). Es besteht bei keinem der Autoren ein Interessenkonflikt.
Ethische Anerkennung
Die Autoren versichern, während des Entstehens der vorliegenden Arbeit, die allgemeingültigen Regeln guter wissenschaftlicher Praxis befolgt zu haben. Da es sich um einen Übersichtsartikel (Literaturrecherche) handelt, kamen keine Tiere zum Einsatz.
Funding
Diese Arbeit wurde durch das Bundesministerium für Bildung und Forschung (BMBF) unter den Projektnummern 01Kl1726A-G als Teil des nationalen Forschungsnetzes zoonotische Infektionskrankheiten finanziert.
Autorenbeitrag
Konzeption der Arbeit: BUB, MR, MG; Datenerhebung, -analyse und -interpretation: BUB, MR, AC, KH, KMS, KB, KS, DF, MRK, SM, CB, AL, SFF, SU, GRM, MG; Manuskriptentwurf: BUB, MR, DF, MRK, SM, CB, AL, GRM, MG; Kritische Revision des Artikels und endgültige Zustimmung der für die Veröffentlichung vorgesehenen Version: BUB, MR, AC, KH, KMS, KB, KS, DF, MRK, SM, CB, AL, SFF, SU, GRM, MG
Korrespondenzadresse
Dr. Benjamin U. Bauer
Klinik für kleine Klauentiere
Stiftung Tierärztliche Hochschule Hannover
Bischofsholer Damm 15
30173 Hannover
Benjamin.Bauer@tiho-hannover.de
Literatur
Achard D, Rodolakis A (2017): Q fever vaccination in ruminants: a critical review. In: Caetano Simoes JC, Ferreira AS, Jorge de Silva G (eds.), The principles and practice of Q fever. Nova Science Publishers Inc, New York, 367–389.
Agerholm JS (2013): Coxiella burnetii associated reproductive disorders in domestic animals – a critical review. Acta Vet Scand 55: 1–11.
Álvarez-Alonso R, Basterretxea M, Barandika JF, Hurtado A, Idiazabal J, Jado I, Beraza X, Montes M, Liendo P, García-Pérez AL (2018): A Q fever outbreak with a high rate of abortions in a dairy goat farm: Coxiella burnetii shedding, environmental contamination and viability. Appl Environ Microbiol 84: e01650-18.
Ammerdorffer A, Schoffelen T, Gresnigt MS, Oosting M, den Brok MH, Abdollahi-Roodsaz S, Kanneganti TD, de Jong DJ, van Deuren M, Roest HJ, Rebel JM, Netea MG, Joosten LA, Sprong T (2015): Recognition of Coxiella burnetii by tolllike receptors and nucleotide-binding oligomerization domainlike receptors. J Infect Dis 211: 978–987.
Andoh M, Zhang G, Russell-Lodrigue KE, Shive HR, Weeks BR, Samuel JE (2007): T cells are essential for bacterial clearance, and gamma interferon, tumor necrosis factor alpha, and B cells are crucial for disease development in Coxiella burnetii infection in mice. Infect Immun 75: 3245–3255.
Angelakis E, Mediannikov O, Jos S-L, Berenger J-M, Parola P, Raoult D (2016): Candidatus Coxiella massiliensis infection. Emerg Infect Dis 22: 285–288.
Angelakis E, Raoult D (2010): Q fever. Vet Microbiol 140: 297–309.
Arricau-Bouvery N, Hauck Y, Bejaoui A, Frangoulidis D, Bodier CC, Souriau A, Meyer H, Neubauer H, Rodolakis A, Vergnaud G (2006): Molecular characterization of Coxiella burnetii isolates by infrequent restriction site-PCR and MLVA typing. BMC Microbiol 6: 38.
Arricau-Bouvery N, Rodolakis A (2005): Is Q fever an emerging or re-emerging zoonosis? Vet Res 36: 327–349.
Arricau-Bouvery N, Souriau A, Bodier C, Dufour P, Rousset E, Rodolakis A (2005): Effect of vaccination with phase I and phase II Coxiella burnetii vaccines in pregnant goats. Vaccine 23: 4392–4402.
Arricau-Bouvery N, Souriau A, Lechopier P, Rodolakis A (2003): Experimental Coxiella burnetii infection in pregnant goats: excretion routes. Vet Res 34: 423–433.
Astobiza I, Barandika JF, Juste RA, Hurtado A, García-Pérez AL (2013): Evaluation of the efficacy of oxytetracycline treatment followed by vaccination against Q fever in a highly infected sheep flock. Vet J 196: 81–85.
Astobiza I, Barandika JF, Ruiz-Fons F, Hurtado A, Povedano I, Juste RA, García-Pérez AL (2011a): Coxiella burnetii shedding and environmental contamination at lambing in two highly naturally-infected dairy sheep flocks after vaccination. Res Vet Sci 91: e58–e63.
Astobiza I, Barandika JF, Ruiz-Fons F, Hurtado A, Povedano I, Juste RA, García-Pérez AL (2011b): A four-year evaluation of the effect of vaccination against Coxiella burnetii on reducing animal infection and environmental contamination in a naturally infected dairy sheep flock. Appl Environ Microbiol 77: 7405–7407.
Avbersek J, Pate M, Skibin A, Ocepek M, Krt B (2019): Management of a Coxiella burnetii-infected sheep flock after an outbreak of Q fever in humans. Turk J Vet Anim Sci 43: 264–270.
Barandika JF, Álvarez-Alonso R, Jado I, Hurtado A, García-Pérez AL (2019): Viable Coxiella burnetii in hard cheeses made with unpasteurized milk. Int J Food Microbiol 303: 42–45.
Beare PA, Gilk SD, Larson CL, Hill J, Stead CM, Omsland A, Cockrell DC, Howe D, Voth DE, Heinzen RA (2011): Dot/
Icm type IVB secretion system requirements for Coxiella burnetii growth in human macrophages. MBio 2: e00175–00111.
Ben Amara A, Ghigo E, Le Priol Y, Lépolard C, Salcedo SP, Lemichez E, Bretelle F, Capo C, Mege J-L (2010): Coxiella burnetii, the agent of Q fever, replicates within trophoblasts and induces a unique transcriptional response. PLoS One 5: e15315.
Benoit M, Barbarat B, Bernard A, Olive D, Mege JL (2008a):
Coxiella burnetii, the agent of Q fever, stimulates an atypical M2 activation program in human macrophages. Eur J Immunol 38: 1065–1070.
Benoit M, Desnues B, Mege JL (2008b): Macrophage polarization in bacterial infections. J Immunol 181: 3733–3739.
Benoit M, Ghigo E, Capo C, Raoult D, Mege J-L (2008c): The uptake of apoptotic cells drives Coxiella burnetii replication and macrophage polarization: a model for Q fever endocarditis. PLoS Pathog 4: e1000066.
Benson W, Brock DW, Mather J (1963): Serologic analysis of a penitentiary group using raw milk from a Q fever infected herd. Public Health Rep 78: 707–710.
Berri M, Crochet D, Santiago S, Rodolakis A (2005): Spread of Coxiella burnetii infection in a flock of sheep after an episode of Q fever. Vet Rec 157: 737–740.
Berri M, Rousset E, Champion J, Russo P, Rodolakis A (2007): Goats may experience reproductive failures and shed Coxiella burnetii at two successive parturitions after a Q fever infection. Res Vet Sci 83: 47–52.
Berri M, Souriau A, Crosby M, Crochet D, Lechopier P, Rodolakis A (2001): Relationships between the shedding of Coxiella burnetii, clinical signs and serological responses of 34 sheep. Vet Rec 148: 502–505.
Berri M, Souriau A, Crosby M, Rodolakis A (2002): Shedding of Coxiella burnetii in ewes in two pregnancies following an episode of Coxiella abortion in a sheep flock. Vet Microbiol 85: 55–60.
BfR (2010): Q-Fieber: Übertragung von Coxiella burnetii durch den Verzehr von Lebensmitteln tierischer Herkunft unwahrscheinlich. Bundesinstitut für Risikobewertung, Berlin, Stellungnahme Nr. 018/2010.
Bisle S, Klingenbeck L, Borges V, Sobotta K, Schulze-Luehrmann J, Menge C, Heydel C, Gomes JP, Lührmann A (2016): The inhibition of the apoptosis pathway by the Coxiella burnetii effector protein CaeA requires the EK repetition motif, but is independent of survivin. Virulence 7: 400–412.
BMEL (2014): Empfehlungen für hygienische Anforderungen an das Halten von Wiederkäuern. Bundesministerium für Ernährung und Landwirtschaft, Bonn.
BMEL (2007): Richtlinie des Bundesministeriums für Ernährung, Landwirtschaft und Verbraucherschutz über Mittel und Verfahren für die Durchführung der Desinfektion bei anzeigepflichtigen Tierseuchen. Bundesministerium für Ernährung Landwirtschaft und Verbraucherschutz, Berlin.
Boettcher J, Schumacher M, Motch B, Mehne D, Alex M, Janowetz B (2017): Feasibility study on Coxiella burnetii phase-specific antibody tests and interferon γ-recall assay in dairy cattle. J Vet Med Res 4: 1106.
Bonkowski K (2016): Analyse der Infektion boviner Epithelzellen mit Coxiella burnetii im in vitro-Modell. Jena, Friedrich-Schiller-Universität Jena, Diss.
Bontje D, Backer J, Hogerwerf L, Roest H, van Roermund H (2016): Analysis of Q fever in Dutch dairy goat herds and assessment of control measures by means of a transmission model. Prev Vet Med 123: 71–89
Botelho-Nevers E, Fournier P-E, Richet H, Fenollar F, Lepidi H, Foucault C, Branchereau A, Piquet P, Maurin M, Raoult D, Diseases I (2007): Coxiella burnetii infection of aortic aneurysms or vascular grafts: report of 30 new cases and evaluation of outcome. Eur J Clin Microbiol Infect Dis 26: 635–640.
Böttcher J, Alex M, Janowetz B (2011a): Application of phase-specific Q fever ELISA in ruminants. Chlamydioses and Q Fever: a zoonotic or livestock problem?, Padua 2011.
Böttcher J, Frangoulidis D, Schumacher M, Janowetz B, Gangl A, Alex M (2013): The impact of Q fever-phase-specific milk serology for the diagnosis of puerperal and chronic milk shedding of C. burnetii in dairy cows. Berl Munch Tierarztl Wochenschr 126: 427–435.
Böttcher J, Niethammer F, Böhm B, Kappe E, Schade B, Turowski V, Bauer B, von Silva-Tarouca B, Domes U, Janowetz B (2019): Ein alternatives Modell der Pathogenese von Q-Fieber-Aborten bei Rindern. Tieraerztl Umschau 74: 216–225.
Böttcher J, Vossen A, Janowetz B, Alex M, Gangl A, Randt A, Meier N (2011b): Insights into the dynamics of endemic Coxiella burnetii infection in cattle by application of phasespecific ELISAs in an infected dairy herd. Vet Microbiol 151: 291–300.
Bradley WP, Boyer MA, Nguyen HT, Birdwell LD, Yu J, Ribeiro JM, Weiss SR, Zamboni DS, Roy CR, Shin S (2016): Primary role for toll-like receptor-driven tumor necrosis factor rather than cytosolic immune detection in restricting Coxiella burnetii phase II replication within mouse macrophages. Infect Immun 84: 998–1015.
Brockmann S (2014): Zoonosen Herausforderungen in endemischen Regionen. Dikussionsforum „Forschung trifft Praxis: Transfer und Herausforderung der Zoonosenforschung“, Berlin 2014.
Brooke RJ, Kretzschmar ME, Mutters NT, Teunis PF (2013): Human dose response relation for airborne exposure to Coxiella burnetii. BMC Infect Dis 13: 488.
Bru J-P, Cazorla C, Choutet P, Fournier P-E, Le Goaster C, Mailles A, Rautureau S, Rousset E (2013): Fièvre Q: recommandations de prise en charge. Haut Conseil de la Santé Publique, Paris.
Capo C, Lindberg FP, Meconi S, Zaffran Y, Tardei G, Brown EJ, Raoult D, Mege JL (1999): Subversion of monocyte functions by Coxiella burnetii: impairment of the cross-talk between alphavbeta3 integrin and CR3. J Immunol 163: 6078–6085.
Carey KL, Newton HJ, Lührmann A, Roy CR (2011): The Coxiella burnetii Dot/Icm system delivers a unique repertoire of type IV effectors into host cells and is required for intracellular replication. PLoS Pathog 7: e1002056.
Carrié P, Barry S, Rousset E, de Cremoux R, Sala C, Calavas D, Perrin JB, Bronner A, Gasqui P, Gilot-Fromont E, Becker CAM, Gache K, Jourdain E (2019): Swab cloths as a tool for revealing environmental contamination by Q fever in ruminant farms. Transbound Emerg Dis 66: 1202–1209.
Cerf O, Condron R (2006): Coxiella burnetii and milk pasteurization: an early application of the precautionary principle? Epidemiol Infect 134: 946–951.
Clark NJ, Soares Magalhães RJ (2018): Airborne geographical dispersal of Q fever from livestock holdings to human communities: a systematic review and critical appraisal of evidence. BMC Infect Dis 18: 218.
Clemente TM, Mulye M, Justis AV, Nallandhighal S, Tran TM, Gilk SD (2018): Coxiella burnetii blocks intracellular interleukin-17 signaling in macrophages. Infect Immun 86: e00532-18.
Coleman SA, Fischer ER, Howe D, Mead DJ, Heinzen RA (2004): Temporal analysis of Coxiella burnetii morphological differentiation. J Bacteriol 186: 7344–7352.
Colombo MI, Gutierrez MG, Romano PS (2006): The two faces of autophagy: Coxiella and Mycobacterium. Autophagy 2: 162–164.
Cunha LD, Ribeiro JM, Fernandes TD, Massis LM, Khoo CA, Moffatt JH, Newton HJ, Roy CR, Zamboni DS (2015): Inhibition of inflammasome activation by Coxiella burnetii type IV secretion system effector IcaA. Nat Commun 6: 10205.
CVUA Stuttgart (2018): Leitfaden zum Q-Fieber Baden-Württemberg. Chemisches- und Veterinäruntersuchungsamt, Stuttgart.
de Cremoux R, Gache K, Rousset E, Sala C, Hosteing S, Nicollet P, Lars F, Guatteo R, Dion F, Calavas D, Bronner A, Perrin J-B, Touratier A (2018): A pilot program for clinical Q fever surveillance as a first step for a standardized differential diagnosis of abortions: organizational lessons applied to goats farms. Small Rumin Res 163: 60–64.
de Cremoux R, Rousset E, Touratier A, Audusseau G, Nicollet P, Ribaud D, David V, Le Pape M (2012): Coxiella burnetii vaginal shedding and antibody responses in dairy goat herds in a context of clinical Q fever outbreaks. FEMS Immunol Med Microbiol 64: 120–122.
DESTATIS (2018a): Rinder, 2 Jahre und älter, Milchkühe. https://www-genesis.destatis.de/genesis/online, (letzter Zugriff 14.05.2019).
DESTATIS (2018b): Weibliche Schafe zur Zucht. https://wwwgenesis.destatis.de/genesis/online, (letzter Zugriff 14.05.2019).
DESTATIS (2016): Ziegen, weiblich. https://www-genesis.destatis.de/genesis/online, 14.05.2019.
Dörner JC (2011): Wirksamkeitsprüfung chemischer Verfahren zur Desinfektion von Coxiella burnetii in kontaminierten Bodenmatrizes. Gießen, Justus-Liebig-Universität Gießen, Diss.
Dupont H, Raoult D, Brouqui P, Janbon F, Peyramond D, Weiller P-J, Chicheportiche C, Nezri M, Poirier R (1992): Epidemiologic features and clinical presentation of acute Q fever in hospitalized patients: 323 French cases. Am J Med 93: 427–434.
Duron O (2015): The IS1111 insertion sequence used for detection of Coxiella burnetii is widespread in Coxiella-like endosymbionts of ticks. FEMS Microbiol Lett 362: fnv132.
Duron O, Sidi-Boumedine K, Rousset E, Moutailler S, Jourdain E (2015): The importance of ticks in Q Fever transmission: what has (and has not) been demonstrated? Trends Parasitol 31: 536–552.
DVG (2019): DVG-Desinfektionsmittelliste für den Tierhaltungsbereich. http://www.desinfektion-dvg.de/index.php?id=2150, (letzter Zugriff 10.06.2019).
Eckart RA, Bisle S, Schulze-Luehrmann J, Wittmann I, Jantsch J, Schmid B, Berens C, Lührmann A (2014): The anti-apoptotic activity of the Coxiella burnetii effector protein AnkG is controlled by p32-dependent trafficking. Infect Immun 82: 2763–2771.
EFSA (2010): Scientific opinion on Q fever. EFSA Journal 8: 1595.
Eibach R, Bothe F, Runge M, Ganter M (2013): Long-term monitoring of a Coxiella burnetti-infected sheep flock after vaccination and antibotic treatment under field conditions. Berl Munch Tierarztl Wochenschr 126: 3–9.
Eldin C, Angelakis E, Renvoisé A, Raoult D (2013): Coxiella burnetii DNA, but not viable bacteria, in dairy products in France. Am J Trop Med Hyg 88: 765–769.
Eldin C, Mélenotte C, Mediannikov O, Ghigo E, Million M, Edouard S, Mege J-L, Maurin M, Raoult D (2017): From Q Fever to Coxiella burnetii Infection: a Paradigm Change. Clin Microbiol Rev 30: 115–190.
EMA (2014): CVMP annual re-assessment report for Coxevac. European Medicines Agency, London.
Fernandes TD, Cunha LD, Ribeiro JM, Massis LM, Lima-Junior DS, Newton HJ, Zamboni DS (2016): Murine alveolar macrophages are highly susceptible to replication of Coxiella burnetii phase II in vitro. Infect Immun 84: 2439–2448.
Fischer S, Göhring-Zwacka E, Kömpf D, Bauer C, Reick D, Pfaff G, Wagner-Wiening C (2016): Labordiagnostische Untersuchung eines großen Q-Fieber-Ausbruchs im Landkreis Heidenheim/Baden-Württemberg. Gesundheitswesen 78: V78.
Fishbein D, Raoult D (1992): A cluster of Coxiella burnetii infections associated with exposure to vaccinated goats and their unpasteurized dairy products. Am J Trop Med Hyg 47: 35–40.
Flannagan RS, Cosío G, Grinstein S (2009): Antimicrobial mechanisms of phagocytes and bacterial evasion strategies. Nat Rev Microbiol 7: 355–366.
Fournier PE, Marrie T, Didier R (1998): Diagnosis of Q fever. J Clin Microbiol 36: 1823–1834.
Frangoulidis D, Walter MC, Antwerpen M, Zimmermann P, Janowetz B, Alex M, Böttcher J, Henning K, Hilbert A, Ganter M, Runge M, Münsterkötter M, Splettstoesser WD, Hanczaruk M (2014): Molecular analysis of Coxiella burnetii in Germany reveals evolution of unique clonal clusters. Int J Med Microbiol 304: 868–876.
Frank C, Höhle M (2011): Just counting sheep? Ecological association of Q fever incidence and ruminant density in Germany. European Scientific Conference on Applied Infectious Disease Epidemiology, Stockholm 2011.
Frentzel H, Menrath A, Tomuzia K, Braeunig J, Appel B (2013): Decontamination of high-risk animal and zoonotic pathogens. Biosecur Bioterror 11 Suppl 1: S102–114.
Friedrich A, Pechstein J, Berens C, Lührmann A (2017): Modulation of host cell apoptotic pathways by intracellular pathogens. Curr Opin Microbiol 35: 88–99.
Gale P, Kelly L, Mearns R, Duggan J, Snary EL (2015): Q fever through consumption of unpasteurised milk and milk products – a risk profile and exposure assessment. J Appl Microbiol 118: 1083–1095.
Ganter M, Bothe F, Eibach R, Runge M (2011): Q fever in small ruminants. Proceedings of the European Conference on Small Ruminant Health Management, Athens 2011.
Ganter M, Bothe F, Runge M, von Keyserlingk M, Braune S, Wichtern B (2009): Ausbruch von Q-Fieber nach Geburtshilfe bei einer Ziege. In: Niedersächsisches Landesamt für Verbraucherschutz und Lebensmittelsicherheit (Hrsg.), Fachbericht zum gesundheitlichen Verbraucherschutz. Niedersächsisches Landesamt für Verbraucherschutz und Lebensmittelsicherheit, Oldenburg, 71–72.
Georgiev M, Alfonso A, Neubauer H, Needham H, Thiéry R, Rodolakis A, Roest HIJ, Stärk KD, Stegeman JA, Vellema P,
van der Hoek W, More SJ (2013): Q fever in humans and farm animals in four European countries, 1982 to 2010. Euro Surveill 18: 20407.
Ghigo E, Honstettre A, Capo C, Gorvel JP, Raoult D, Mege JL(2004): Link between impaired maturation of phagosomes and defective Coxiella burnetii killing in patients with chronic Q fever. J Infect Dis 190: 1767–1772.
Gilsdorf A, Kroh C, Grimm S, Jensen E (2008): Large Q fever outbreak due to sheep farming near residential areas, Germany, 2005. Epidemiol Infect 136: 1084–1087.
Glazunova O, Roux V, Freylikman O, Sekeyova Z, Fournous G, Tyczka J, Tokarevich N, Kovacava E, Marrie TJ, Raoult D (2005): Coxiella burnetii genotyping. Emerg Infect Dis 11: 1211–1217.
González-Barrio D, Jado I, Fernández-de-Mera IG, del Rocio Fernández-Santos M, Rodríguez-Vargas M, García-Amil C, Beltrán-Beck B, Anda P, Ruiz-Fons F (2016): Genotypes of Coxiella burnetii in wildlife: disentangling the molecular epidemiology of a multi-host pathogen. Env Microbiol Rep 8: 708–714.
Gottlieb Y, Lalzar I, Klasson L (2015): Distinctive genome reduction rates revealed by genomic analyses of two Coxiella-like
endosymbionts in ticks. Genome Biol Evol 7: 1779–1796.
Guatteo R, Beaudeau F, Joly A, Seegers H (2007): Coxiella burnetii shedding by dairy cows. Vet Res 38: 849–860.
Hamann HP, Volmer R, Wimmershof N, Ballmann G, Zschöck M (2009): Q-Fieber-Impfung in zwei Schafhaltungen nach gehäuftem Auftreten humaner Erkrankungen. Tieraerztl Umschau 64: 188–190.
Hatchette T, Campbell N, Hudson R, Raoult D, Marrie TJ (2003): Natural history of Q fever in goats. Vector-Borne Zoonot 3: 11–15.
Hawker J, Ayres J, Blair I, Evans MR, Smith D, Smith E, Burge P, Carpenter M, Caul E, Coupland B (1998): A large outbreak of Q fever in the West Midlands: windborne spread into a metropolitan area? Commun Dis Public Health 1: 180–187.
Hazlett MJ, McDowall R, DeLay J, Stalker M, McEwen B, van Dreumel T, Spinato M, Binnington B, Slavic D, Carman S, Cai HY (2013): A prospective study of sheep and goat abortion using real-time polymerase chain reaction and cut point estimation shows Coxiella burnetii and Chlamydophila abortus infection concurrently with other major pathogens. J Vet Diagn Invest 25: 359–368.
Hechemy KE (2012): History and prospects of Coxiella burnetii research. In: Toman R, Heinzen R, Samuel JE, Mege J (.), Coxiella burnetii: recent advances and new perspectives in research in the Q fever bacterium. Springer, Dordrecht, 1–11.
Hellenbrand W, Breuer T, Petersen L (2001): Changing epidemiology of Q fever in Germany, 1947–1999. Emerg Infect Dis 7: 789–796.
Hermans MH, Huijsmans CR, Schellekens JJ, Savelkoul PH, Wever PC (2011): Coxiella burnetii DNA in goat milk after vaccination with Coxevac®. Vaccine 29: 2653–2656.
Hilbert A, Blaha I, Hensler E, Reith P, Henning K, Conraths FJ, Miller T (2014): Aspekte seroepidemiologischer Untersuchungen zum Q-Fieber in nicht geimpften Rinderbeständen. Berl Munch Tierarztl Wochenschr 127: 149–157.
Hilbert A, Reith P, Brockmann SO, Tyczka J, Fischer SF, Piechotowski I, Wagner-Wiening C, Winter CH, Bendak J, Meier C (2011): Epidemiologische Untersuchungen zu zwei Q-Fieber-Ausbrüchen in einer Gemeinde Baden-Württembergs in den Jahren 2008 und 2009. Berl Munch Tierarztl Wochenschr 124: 295–302.
Hilbert A, Schmoock G, Lenzko H, Moog U, Diller R, Fröhlich A, Hoffmann L, Horner S, Elschner M, Tomasso H, Henning K, Neubauer H, Sprague LD (2012): Prevalence of Coxiella burnetii in clinically healthy German sheep flocks. BMC Res Notes 5: 152
Hildebrandt A, Straube E, Neubauer H, Schmoock G (2011): Coxiella burnetii and coinfections in Ixodes ricinus ticks in central Germany. Vector-Borne Zoonot 11: 1205-1207.
Hill J, Samuel JE (2011): Coxiella burnetii acid phosphatase inhibits the release of reactive oxygen intermediates in polymorphonuclear leukocytes. Infect Immun 79: 414–420.
Hogerwerf L, Courcoul A, Klinkenberg D, Beaudeau F, Vergu E, Nielen M (2013): Dairy goat demography and Q fever infection dynamics. Vet Res 44: 28.
Honstettre A, Ghigo E, Moynault A, Capo C, Toman R, Akira S, Takeuchi O, Lepidi H, Raoult D, Mege JL (2004): Lipopolysaccharide from Coxiella burnetii is involved in bacterial phagocytosis, filamentous actin reorganization, and inflammatory responses through toll-like receptor 4. J Immunol 172: 3695–3703.
Hotta A, Zhang GQ, Andoh M, Yamaguchi T, Fukushi H, Hirai K (2004): Use of monoclonal antibodies for analyses of Coxiella burnetii major antigens. J Vet Med Sci 66: 1289–1291.
Howe D, Shannon JG, Winfree S, Dorward DW, Heinzen RA (2010): Coxiella burnetii phase I and II variants replicate with similar kinetics in degradative phagolysosome-like compartments of human macrophages. Infect Immun 78: 3465–3474.
IfSG (2019): Infektionsschutzgesetz vom 20. Juli 2000 (BGBI. I S. 1045), das zuletzt durch Artikel 14a der Verordnung vom 6. Mai 2019 (BGBI. I S. 646) geändert worden ist.
Islam A, Lockhart M, Stenos J, Graves S (2013): The attenuated Nine Mile phase II clone 4/RSA439 strain of Coxiella burnetii is highly virulent for severe combined immunodeficient (SCID) mice. Am J Trop Med Hyg 89: 800–803.
Janowetz B, Bauer B, Turowski V, Alex M, Domes U, Böttcher J (2018): Die Impfung und Überwachung von Coxevac®-geimpften Zutretern in einer infizierten Schafherde über 5 Jahre. 5. Jenaer Q-Fieber-Workshop, Jena 2018.
Joulié A, Laroucau K, Bailly X, Prigent M, Gasqui P, Lepetitcolin E, Blanchard B, Rousset E, Sidi-Boumedine K, Jourdain E (2015): Circulation of Coxiella burnetii in a naturally infected flock of dairy sheep: shedding dynamics, environmental contamination, and genotype diversity. Appl Environ Microbiol 81: 7253–7260.
Joulié A, Rousset E, Gasqui P, Lepetitcolin E, Leblond A, Sidi-Boumedine K, Jourdain E (2017): Coxiella burnetii circulation in a naturally infected flock of sheep: individual follow-up of antibodies in serum and milk. Appl Environ Microbiol 83: e00222-17.
Jourdain E, Duron O, Barry S, González-Acuña D, Sidi-Boumedine K (2015): Molecular methods routinely used to detect Coxiella burnetii in ticks cross-react with Coxiella-like bacteria. Infect Ecol Epidemiol 5: 29230.
Kampschreur LM, Hagenaars JC, Wielders CC, Elsman P, Lestrade PJ, Koning OH, Oosterheert JJ, Renders NH, Wever PC (2013): Screening for Coxiella burnetii seroprevalence in chronic Q fever high-risk groups reveals the magnitude of the Dutch Q fever outbreak. Epidemiol Infect 141: 847–851.
Kersh GJ, Fitzpatrick KA, Self JS, Priestley RA, Kelly AJ, Lash RR, Marsden-Haug N, Nett RJ, Bjork A, Massung RF, Anderson AD (2013): Presence and persistence of Coxiella burnetii in the environment of goat farms associated with a Q fever outbreak. Appl Environ Microbiol 79: 1697–1703.
Kersh GJ, Wolfe TM, Fitzpatrick KA, Candee AJ, Oliver LD, Patterson NE, Self JS, Priestley RA, Loftis AD, Massung RF (2010): Presence of Coxiella burnetii DNA in the environment of the United States, 2006 to 2008. Appl Environ Microbiol 76: 4469–4475.
Khavkin T, Tabibzadeh SS (1988): Histologic, immunofluorescence, and electron microscopic study of infectious process in mouse lung after intranasal challenge with Coxiella burnetii. Infect Immun 56: 1792–1799.
Kimmig P (2010): Q-Fieber – Eine Infektion mit komplexer Epidemiologie. In: Gusenleitner F, Aescht E (Hrsg.), Krank durch Arthropoden – Denisia 30. Land Oberösterreich, Biologiezentrum/ Oberösterreichische Landesmuseen, Linz, 593–604.
King LA, Goirand L, Tissot-Dupont H, Giunta B, Giraud C, Colardelle C, Duquesne V, Rousset E, Aubert M, Thiery R (2011): Outbreak of Q fever, Florac, southern France, spring 2007. Vector-Borne Zoonot 11: 341–347.
Kishimoto RA, Rozmiarek H, Larson EW (1978): Experimental Q fever infection in congenitally athymic nude mice. Infect Immun 22: 69–71.
Kittelberger R, Mars J, Wibberley G, Sting R, Henning K, Horner GW, Garnett KM, Hannah MJ, Jenner JA, Piggott CJ, O’Keefe JS (2009): Comparison of the Q-fever complement fixation test and two commercial enzyme-linked immunosorbent assays for the detection of serum antibodies against Coxiella burnetti (Q-fever) in ruminants : recommendations for use of serological tests on imported animals in New Zealand. NZ Vet J 57: 262–268.
Klee SR, Tyczka J, Ellerbrok H, Franz T, Linke S, Baljer G, Appel B (2006): Highly sensitive real-time PCR for specific detection and quantification of Coxiella burnetii. BMC Microbiol 6: 2.
Klingenbeck L, Eckart RA, Berens C, Lührmann A (2013): The Coxiella burnetii type IV secretion system substrate CaeB inhibits intrinsic apoptosis at the mitochondrial level. Cell Microbiol 15: 675–687.
Landwirtschaftskammer Nordrhein-Westfalen (2014): Beihilferichtlinien für Schafe. https://www.landwirtschaftskammer.de/landwirtschaft/tierseuchenkasse/le…, 21.04.2019.
Larson CL, Beare PA, Howe D, Heinzen RA (2013): Coxiella burnetii effector protein subverts clathrin-mediated vesicular trafficking for pathogen vacuole biogenesis. Proc Natl Acad Sci U S A 110: E4770-9.
Latomanski EA, Newton P, Khoo CA, Newton HJ (2016): The effector Cig57 hijacks FCHO-mediated vesicular trafficking to facilitate intracellular replication of Coxiella burnetii. PLoS Pathog 12: e1006101.
Lehner S, Lohan K, Dieckhoff H-J, Gerdes U (2017): Erfahrungen von Tierhaltern in niedersächsischen Milchkuhbetrieben mit der Impfung gegen Q-Fieber. Tierarztl Prax G N 45: 141–149.
Liebisch A (1977): Das Q-Fieber als Naturherdinfektion in Süddeutschland. Bundesgesundheitsbl 20: 185–191.
Limonard GJ, Thijsen SF, Bossink AW, Asscheman A, Bouwman JJ (2012): Developing a new clinical tool for diagnosing chronic Q fever: the Coxiella ELISPOT. FEMS Immunol Med Microbiol 64: 57–60
Lucchese L, Capello K, Barberio A, Zuliani F, Stegeman A, Ceglie L, Guerrini E, Marangon S, Natale A (2015): IFAT and ELISA phase I/phase II as tools for the identification of Q fever chronic milk shedders in cattle. Vet Microbiol 179: 102–108.
Lührmann A, Newton HJ, Bonazzi M (2017): Beginning to understand the role of the type IV secretion system effector proteins in Coxiella burnetii pathogenesis. In: Backert S, Grohmann E (eds.), Type IV Secretion in Gram-Negative and Gram-Positive Bacteria. Springer International Publishing, Cham, 243–268.
Lührmann A, Nogueira CV, Carey KL, Roy CR (2010): Inhibition of pathogen-induced apoptosis by a Coxiella burnetii type IV effector protein. Proc Natl Acad Sci U S A 107: 18997–19001.
Lührmann A, Roy CR (2007): Coxiella burnetii inhibits activation of host cell apoptosis through a mechanism that involves preventing cytochrome c release from mitochondria. Infect Immun 75: 5282–5289.
Martinez E, Allombert J, Cantet F, Lakhani A, Yandrapalli N, Neyret A, Norville IH, Favard C, Muriaux D, Bonazzi M (2016): Coxiella burnetii effector CvpB modulates phosphoinositide metabolism for optimal vacuole development. Proc Natl Acad Sci U S A 113: E3260-69.
Martinez E, Cantet F, Fava L, Norville I, Bonazzi M (2014): Identification of OmpA, a Coxiella burnetii protein involved in host cell invasion, by multi-phenotypic high-content screening. PLoS Pathog 10: e1004013.
Martinov SP (2007): Contemporary state of the problem Q fever in Bulgaria. Biotechnol Biotechnol Equip 21: 353–361.
Massung RF, Cutler SJ, Frangoulidis D (2012): Molecular typing of Coxiella burnetii (Q Fever). In: Toman R, Heinzen R, Samuel JE, Mege J (eds.), Coxiella burnetii: recent advances and new perspectives in research in the Q fever bacterium. Springer, Dordrecht, 382–396.
Maurin M, Raoult DF (1999): Q fever. Clin Microbiol Rev 12: 518–553.
McCaul T, Williams J (1981): Developmental cycle of Coxiella burnetii: structure and morphogenesis of vegetative and sporogenic differentiations. J Bacteriol 147: 1063–1076.
Meghari S, Berruyer C, Lepidi H, Galland F, Naquet P, Mege J-L (2007): Vanin-1 controls granuloma formation and macrophage polarization in Coxiella burnetii infection. Eur J Immunol 37: 24–32.
Mehraj V, Textoris J, Ben Amara A, Ghigo E, Raoult D, Capo C, Mege J-L (2013): Monocyte responses in the context of Q fever: from a static polarized model to a kinetic model of activation. J Infect Dis 208: 942–951.
Melenotte C, Protopopescu C, Million M, Edouard S, Carrieri MP, Eldin C, Angelakis E, Djossou F, Bardin N, Fournier PE, Mege JL, Raoult D (2018): Clinical features and complications of Coxiella burnetii infections from the French national reference center for Q fever. JAMA Netw Open 1: e181580.
Million M, Thuny F, Richet H, Raoult D (2010): Long-term outcome of Q fever endocarditis: a 26-year personal survey. Lancet Infect Dis 10: 527–535.
Minnick MF, Raghavan R (2012): Developmental biology of Coxiella burnetii. In: Toman R, Heinzen RA, Samuel JE, Mege J-L (eds.), Coxiella burnetii: recent advances and new perspectives in research of the Q Fever bacterium. Springer, Dordrecht, 231–248.
Moeller RB (2012): Disorders of sheep and goats. In: Njaa BL (ed.), Kirkbride’s diagnosis of abortion and neonatal loss in animals. Wiley-Blackwell, West Sussex.
Moog U, Henning K, Horner S (2018): Q-Fieber Bekämpfung bei Schafen und Ziegen in Thüringen. 5. Jenaer Q-Fieber-Workshop, Jena 2018.
Moog U, Henning K, Horner S (2016): Q fever - vaccination and surveillance in Thuringia. Annual European College of Small Ruminant Health Management Conference, Freiburg im Breisgau 2016.
Mori M, Roest H-J (2018): Farming, Q fever and public health: agricultural practices and beyond. Arch Public Health 76: 2.
Morroy G, Keijmel SP, Delsing CE, Bleijenberg G, Langendam M, Timen A, Bleeker-Rovers CP (2016): Fatigue following acute Q-fever: a systematic literature review. PLoS One 11: e0155884.
Mosser DM, Edwards JP (2008): Exploring the full spectrum of macrophage activation. Nat Rev Immunol 8: 958–969.
Muleme M, Stenos J, Vincent G, Campbell A, Graves S, Warner S, Devlin JM, Nguyen C, Stevenson MA, Wilks CR, Firestone SM (2016): Bayesian validation of the indirect immunofluorescence assay and its superiority to the enzyme-linked immunosorbent assay and the complement fixation test for detecting antibodies against Coxiella burnetii in goat serum. Clin Vaccine Immunol 23: 507–514.
Muleme M, Stenos J, Vincent G, Wilks CR, Devlin JM, Campbell A, Cameron A, Stevenson MA, Graves S, Firestone SM (2017): Peripartum dynamics of Coxiella burnetii infections in intensively managed dairy goats associated with a Q fever outbreak in Australia. Prev Vet Med 139: 58–66.
Natale A, Bucci G, Capello K, Barberio A, Tavella A, Nardelli S, Marangon S, Ceglie L (2012): Old and new diagnostic approaches for Q fever diagnosis: correlation among serological (CFT, ELISA) and molecular analyses. Comp Immunol Microbiol Infect Dis 35: 375–379.
Newton HJ, Kohler LJ, McDonough JA, Temoche-Diaz M, Crabill E, Hartland EL, Roy CR (2014): A screen of Coxiella burnetii mutants reveals important roles for Dot/Icm effectors and host autophagy in vacuole biogenesis. PLoS Pathog 10: e1004286.
OIE (2018): Q fever. In: Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. OIE (World Organisation for Animal Health), Paris, 560–576
Palmer N, Kierstead M, Key D, Williams J, Peacock M, Vellend H (1983): Placentitis and abortion in goats and sheep in Ontario caused by Coxiella burnetii. Can Vet J 24: 60–61.
Pexara A, Solomakos N, Govaris A (2018): Q fever and prevalence of Coxiella burnetii in milk. Trends Food Sci Tech 71: 65–72.
Plummer PJ, McClure JT, Menzies P, Morley PS, Van den Brom R, Van Metre DC (2018): Management of Coxiella burnetii infection in livestock populations and the associated zoonotic risk: a consensus statement. J Vet Intern Med 32: 1481–1494.
Pluta S, Hartelt K, Oehme R, Mackenstedt U, Kimmig P (2010): Prevalence of Coxiella burnetii and Rickettsia spp. in ticks and rodents in southern Germany. Ticks Tick-Borne Dis 1: 145–147.
Porten K, Rissland J, Tigges A, Broll S, Hopp W, Lunemann M, van Treeck U, Kimmig P, Brockmann SO, Wagner-Wiening C, Hellenbrand W, Buchholz U (2006): A super-spreading ewe infects hundreds with Q fever at a farmers’ market in Germany. BMC Infect Dis 6: 147.
Priestley R (2007): Decontamination issues with Coxiella burnetii: an evaluation of currently available disinfectants. American Society of Rickettsiology 21st Meeting, Colorado Springs 2007.
Ramstead AG, Robison A, Blackwell A, Jerome M, Freedman B, Lubick KJ, Hedges JF, Jutila MA (2016): Roles of toll-like receptor 2 (TLR2), TLR4, and MyD88 during pulmonary Coxiella burnetii infection. Infect Immun 84: 940–949.
Raoult D, Tissot-Dupont H, Foucault C, Gouvernet J, Fournier PE, Bernit E, Stein A, Nesri M, Harle JR, Weiller PJ (2000): Q fever 1985-1998. Clinical and epidemiologic features of 1,383 infections. Medicine 79: 109–123.
Read AJ, Erickson S, Harmsen AG (2010): Role of CD4+ and CD8+ T cells in clearance of primary pulmonary infection with oxiella burnetii. Infect Immun 78: 3019-3026.
Reintjes R, Hellenbrand W, Düsterhaus A (2000): Q-Fieber-Ausbruch in Dortmund im Sommer 1999. Gesundheitswesen 62: 609–614.
RKI (2002): Aufklärung eines Q-Fieber-Ausbruchs durch Erkrankungen in einem Film-Team. Epid Bull 37: 316–317.
RKI (2001): Ein Q-Fieber-Ausbruch im Hochsauerland und Nordhessen. Epid Bull 26: 187–189.
RKI (2004): Ermittlungen zu einem Q-Fieber-Ausbruch in einer Großfamilie. Epid Bull 26: 205–207.
RKI (2017): Liste der vom Robert Koch-Institut geprüften und anerkannten Desinfektionsmittel und -verfahren. Bundesgesundheitsbl 60: 1274–1297.
RKI (2008): Q-Fieber: Vermehrtes Auftreten im Frühjahr 2008. Epid Bull 25: 199–203.
RKI (2012): RKI-Ratgeber: Q-Fieber. https://www.rki.de/DE/Content/Infekt/EpidBull/Merkblaetter/Ratgeber_Q-F…, 15.05.2019.
Rodolakis A (2009): Q fever in dairy animals. Ann N Y Acad Sci 1166: 90–93.
Rodolakis A, Berri M, Hechard C, Caudron C, Souriau A, Bodier C, Blanchard B, Camuset P, Devillechaise P, Natorp J (2007): Comparison of Coxiella burnetii shedding in milk of dairy bovine, caprine, and ovine herds. J Dairy Sci 90: 5352–5360.
Roest H, Post J, van Gelderen B, van Zijderveld FG, Rebel JM (2013): Q fever in pregnant goats: humoral and cellular immune responses. Vet Res 44: 67.
Roest H, van Gelderen B, Dinkla A, Frangoulidis D, van Zijderveld F, Rebel J, van Keulen L (2012): Q fever in pregnant goats: pathogenesis and excretion of Coxiella burnetii. PLoS One 7: e48949.
Rogalski W (1985): Chemical modification of the tetracyclines. In: Hlavka JJ, Boothe JH (eds.), The Tetracyclines. Springer, Berlin, 179–316.
Rolain JM, Lambert F, Raoult D (2005): Activity of telithromycin against thirteen new isolates of C. burnetii including three resistant to doxycycline. Ann N Y Acad Sci 1063: 252–256.
Rouli L, Rolain JM, El Filali A, Robert C, Raoult D (2012): Genome sequence of Coxiella burnetii 109, a doxycycline-resistant clinical isolate. J Bacteriol 194: 6939.
Rousset E, Berri M, Durand B, Dufour P, Prigent M, Delcroix T, Touratier A, Rodolakis A (2009a): Coxiella burnetii shedding routes and antibody response after outbreaks of Q feverinduced abortion in dairy goat herds. Appl Environ Microbiol 75: 428–433.
Rousset E, Durand B, Berri M, Dufour P, Prigent M, Russo P, Delcroix T, Touratier A, Rodolakis A, Aubert M (2007): Comparative diagnostic potential of three serological tests for abortive Q fever in goat herds. Vet Microbiol 124: 286–297.
Rousset E, Durand B, Champion JL, Prigent M, Dufour P, Forfait C, Marois M, Gasnier T, Duquesne V, Thiery R, Aubert MF (2009b): Efficiency of a phase 1 vaccine for the reduction of vaginal Coxiella burnetii shedding in a clinically affected goat herd. Clin Microbiol Infect 15 Suppl 2: 188–189.
Rubel F, Brugger K, Monazahian M, Habedank B, Dautel H, Leverenz S, Kahl O (2014): The first German map of georeferenced ixodid tick locations. Parasite Vector 7: 477.
Ruiz-Fons F, Astobiza I, Barandika JF, Juste RA, Hurtado A, Garcia-Perez AL (2011): Measuring antibody levels in bulktank milk as an epidemiological tool to search for the status of Coxiella burnetii in dairy sheep. Epidemiol Infect 139: 1631–1636.
Runge M, Binder A, Schotte U, Ganter M (2012): Investigations concerning the prevalence of Coxiella burnetii and Chlamydia abortus in sheep in correlation with management systems and abortion rate in Lower Saxony in 2004. Berl Munch Tierarztl Wochenschr 125: 10–15.
Sánchez J, Souriau A, Buendía AJ, Arricau-Bouvery N, Martínez CM, Salinas J, Rodolakis A, Navarro JA (2006): Experimental Coxiella burnetii infection in pregnant goats: a histopathological and immunohistochemical study. J Comp Pathol 135: 108–115.
Schäfer W, Eckart RA, Schmid B, Cagköylü H, Hof K, Muller YA, Amin B, Lührmann A (2017): Nuclear trafficking of the anti-apoptotic Coxiella burnetii effector protein AnkG requires binding to p32 and Importin-α1. Cell Microbiol 19: e12634.
Schimmer B, Luttikholt S, Hautvast JLA, Graat EAM, Vellema P, van Duynhoven YTHP (2011): Seroprevalence and risk factors of Q fever in goats on commercial dairy goat farms in the Netherlands, 2009-2010. BMC Vet Res 7: 81.
Schliesser T (1991): Epidemiology and importance of Q fever in German livestock. / Zur Epidemiologie und Bedeutung des Q-Fiebers bei Tieren. Wien Tieraerztl Monat 78: 7–12.
Schmeer N (1985): Enzyme-linked immunosorbent assay (ELISA) for the demonstration of IgG1, IgG2 and IgM antibodies in bovine Q fever infection. Zentralbl Bakteriol Mikrobiol Hyg A 259: 20–34.
Schoenlaub L, Cherla R, Zhang Y, Zhang G (2016): Coxiella burnetii avirulent Nine Mile phase II induces caspase-1 dependent pyroptosis in murine peritoneal B1a B cells. Infect Immun 84: 3638–3654.
Schoffelen T, Ammerdorffer A, Hagenaars JC, Bleeker-Rovers CP, Wegdam-Blans MC, Wever PC, Joosten LA, van der Meer JW, Sprong T, Netea MG, van Deuren M, van de Vosse E (2015): Genetic variation in pattern recognition receptors and adaptor proteins associated with development of chronic Q Fever. J Infect Dis 212: 818–829.
Schüle A (2008): Seroepidemiologische Studie zum Nachweis von Antikörpern gegen Coxiella burnetii und Chlamydophila psittaci bei Zootieren und den betreuenden Personen. Berlin, Freien Universität Berlin, Diss.
Schulz J, Runge M, Schröder C, Ganter M, Hartung J (2005): Detection of Coxiella burnetii in the air of a sheep barn during shearing. Deut Tieraertzl Woch 112: 470–472.
Schulze-Luehrmann J, Eckart RA, Ölke M, Saftig P, Liebler-Tenorio E, Lührmann A (2016): LAMP proteins account for the maturation delay during the establishment of the Coxiella burnetii-containing vacuole. Cell Microbiol 18: 181–194.
Selbitz H-J, Ganter M (2018): Bakterielle Zoonosen: Q-Fieber. In: Bostedt H, Ganter M, Hiepe T (Hrsg.), Lehrbuch der Schaf- und Ziegenkrankheiten. Thieme, Stuttgart, 406–407.
Serbezov VS, Kazar J, Novkirishki V, Gatcheva N, Kovacova E, Voynova V (1999): Q fever in Bulgaria and Slovakia. Emerg Infect Dis 5: 388-394.
Seshadri R, Paulsen IT, Eisen JA, Read TD, Nelson KE, Nelson WC, Ward NL, Tettelin H, Davidsen TM, Beanan MJ (2003): Complete genome sequence of the Q-fever pathogen Coxiella burnetii. Proc Natl Acad Sci U S A 100: 5455–5460.
Shannon JG, Howe D, Heinzen RA (2005): Virulent Coxiella burnetii does not activate human dendritic cells: role of lipopolysaccharide as a shielding molecule. Proc Natl Acad Sci U S A 102: 8722–8727.
Shivaprasad H, Cadenas M, Diab S, Nordhausen R, Bradway D, Crespo R, Breitschwerdt E (2008): Coxiella-like infection in psittacines and a toucan. Avian Dis 52: 426–432.
Sidi-Boumedine K, Rousset E, Henning K, Ziller M, Niemczuck K, Roest H, Thiéry R (2010): Development of harmonised schemes for the monitoring and reporting of Q-fever in animals in the European Union. Scientific Report submitted to EFSA 7: 48E.
Siemsen DW, Kirpotina LN, Jutila MA, Quinn MT (2009): Inhibition of the human neutrophil NADPH oxidase by Coxiella burnetii. Microbes Infect 11: 671–679.
Signs KA, Stobierski MG, Gandhi TN (2012): Q fever cluster among raw milk drinkers in Michigan, 2011. Clin Infect Dis 55: 1387–1389.
Sobotta K, Hillarius K, Mager M, Kerner K, Heydel C, Menge C (2016): Coxiella burnetii infects primary bovine macrophages and limits their host cell response. Infect Immun 84: 1722–1734.
Špitalská E, Kocianová E (2003): Detection of Coxiella burnetii in ticks collected in Slovakia and Hungary. Eur J Epidemiol 18: 263–266.
Sprong H, Tijsse-Klasen E, Langelaar M, De Bruin A, Fonville M, Gassner F, Takken W, Van Wieren S, Nijhof A, Jongejan F, Maassen CBM, Scholte E-J, Hovius JW, Emil Hovius K, Špitalská E, Van Duynhoven YT (2012): Prevalence of Coxiella burnetii in ticks after a large outbreak of Q fever. Zoonoses Public Health 59: 69–75.
Stein A, Saunders N, Taylor A, Raoult D (1993): Phylogenic homogeneity of Coxiella burnetii strains as determinated by 16S ribosomal RNA sequencing. FEMS Microbiol Lett 113: 339–344.
Steinman RM, Nussenzweig MC (2002): Avoiding horror autotoxicus: the importance of dendritic cells in peripheral T cell tolerance. Proc Natl Acad Sci U S A 99: 351–358.
StIKoVet (2018): Stellungnahme zur Umwidmung von immunologischen Tierarzneimitteln. Ständige Impfkommission Veterinärmedizin (StIKo Vet) am Friedrich-Loeffler-Institut, Greifswald - Insel Riems.
Sting R, Breitling N, Oehme R, Kimmig P (2004): Untersuchungen zum Vorkommen von Coxiella burnetii bei Schafen und Zecken der Gattung Dermacentor in Baden-Wuerttemberg. Deut Tieraertzl Woch 111: 390–394.
Sting R, Molz K, Philipp W, Bothe F, Runge M, Ganter M (2013): Quantitative real-time PCR and phase specific serology are mutually supportive in Q fever diagnostics in goats. Vet Microbiol 167: 600–608.
Tan C, Owens L (2000): Infectivity, transmission and 16S rRNA sequencing of a rickettsia, Coxiella cheraxi sp. nov., from the freshwater crayfish Cherax quadricarinatus. Dis Aquat Organ 41: 115–122.
Taurel AF, Guatteo R, Joly A, Beaudeau F (2012): Effectiveness of vaccination and antibiotics to control Coxiella burnetii shedding around calving in dairy cows. Vet Microbiol 159: 432–437.
Theegarten D, Garnier Y, Ebsen M, Peters M, Hotzel H, Henning K (2004): Q-Fieber Gruppeninfektion im Rahmen von Versuchen zur fetalen Asphyxie im Schafmodell. Pneumologie 58: VPPRS_11.
TierGesG (2019): Tiergesundheitsgesetz in der Fassung der Bekanntmachung vom 21. November 2018 (BGBl. I S. 1938), das durch Artikel 100 des Gesetzes vom 20. November 2019 (BGBl. I S. 1626) geändert worden ist.
Tierseuchenkasse Baden-Württemberg (2018): Satzung der Tierseuchenkasse Baden-Württemberg über die Gewährung von Beihilfen und sonstigen Leistungen (Leistungssatzung) vom 19. Dezember 2014, geändert durch Satzungen vom 11. Februar 2015 und 23. November 2018. Tierseuchenkasse Baden-Württemberg, Stuttgart.
Tierseuchenkasse Hessen (2016): Richtlinien der Hessischen Tierseuchenkasse über die Gewährung von Leistungen (Beihilferichtlinien). Tierseuchenkasse Hessen, Wiesbaden.
Tierseuchenkasse Rheinland-Pfalz (2017): Satzung der Tierseuchenkasse Rheinland-Pfalz über die Gewährung von Beihilfen und Leistungen (Beihilfesatzung). Tierseuchenkasse Rheinland-Pfalz, Bad Kreuznach.
Tierseuchenkasse Sachsen (2015): Neufassung des Programms der Sächsischen Tierseuchenkasse zur Diagnostik und Bekämpfung der Q-Fieber-Infektion bei Rindern, Schafen und Ziegen. Tierseuchenkasse Sachsen, Dresden.
Tierseuchenkasse Thüringen (2019): Satzung über Beihilfen der Thüringer Tierseuchenkasse (Beihilfesatzung) vom 17. November 2015, zuletzt geändert und beschlossen am 22. Februar 2019. Tierseuchenkasse Thüringen, Jena.
Tissot-Dupont H, Amadei M-A, Nezri M, Raoult D (2004): Wind in November, Q fever in December. Emerg Infect Dis 10: 1264–1269.
Tissot-Dupont H, Torres S, Nezri M, Raoult D (1999): Hyperendemic focus of Q fever related to sheep and wind. Am J Epidemiol 150: 67–74.
TKrMeldpflV (2015): Verordnung über meldepflichtige Tierkrankheiten in der Fassung der Bekanntmachung vom 11. Februar 2011 (BGBI. I S. 252), die zuletzt durch Artikel 391 der Verordnung vom 31. August 2015 (BGBI. I S. 1474) geändert worden ist.
Todkill D, Fowler T, Hawker JI (2018): Estimating the incubation period of acute Q fever, a systematic review. Epidemiol Infect 146: 665–672.
USAMRIID (2014): Q-Fever. In: Withers MR (ed.), USAMRIID´s Medical Management of Biological Casualties Handbook. USAMRIID, Fort Detrick.
van den Brom R, Roest HJ, de Bruin A, Dercksen D, Santman-Berends I, van der Hoek W, Dinkla A, Vellema J, Vellema P (2015a): A probably minor role for land-applied goat manure in the transmission of Coxiella burnetii to humans in the 2007-2010 Dutch Q fever outbreak. PLoS One 10: e0121355.
van den Brom R, Santman-Berends I, Luttikholt S, Moll L, Van Engelen E, Vellema P (2015b): Bulk tank milk surveillance as a measure to detect Coxiella burnetii shedding dairy goat herds in the Netherlands between 2009 and 2014. J Dairy Sci 98: 3814–3825.
van den Brom R, Van E, Vos J, Luttikholt SJM, Moll L, Roest HIJ, van der Heijden HMJF, Vellema P (2013): Detection of Coxiella burnetii in the bulk tank milk from a farm with vaccinated goats, by using a specific PCR technique. Small Rumin Res 110: 150–154.
van den Brom R, van Engelen E, Luttikholt S, Moll L, van Maanen K, Vellema P (2012): Coxiella burnetii in bulk tank milk samples from dairy goat and dairy sheep farms in The Netherlands in 2008. Vet Rec 170: 310.
van den Brom R, van Engelen E, Roest HIJ, van der Hoek W, Vellema P (2015c): Coxiella burnetii infections in sheep or goats: an opinionated review. Vet Microbiol 181: 119–129.
van den Brom R, Vellema P (2009): Q fever outbreaks in small ruminants and people in the Netherlands. Small Rumin Res 86: 74–79.
van der Hoek W, Dijkstra F, Schimmer B, Schneeberger P, Vellema P, Wijkmans C, Ter Schegget R, Hackert V, van Duynhoven Y (2010): Q fever in the Netherlands: an update on the epidemiology and control measures. Euro Surveill 15: 19520.
van der Hoek W, Hogema BM, Dijkstra F, Rietveld A, Wijkmans CJ, Schneeberger PM, Zaaijer HL (2012): Relation between Q fever notifications and Coxiella burnetii infections during the 2009 outbreak in The Netherlands. Euro Surveill 17: 20058.
van Leuken JPG, Swart AN, Brandsma J, Terink W, van de Kassteele J, Droogers P, Sauter F, Havelaar AH, van der Hoek W (2016): Human Q fever incidence is associated to spatiotemporal environmental conditions. One Health 2: 77–87.
van Schaik EJ, Chen C, Mertens K, Weber MM, Samuel JE (2013): Molecular pathogenesis of the obligate intracellular bacterium Coxiella burnetii. Nat Rev Microbiol 11: 561–573.
Vellema P, Moll L, van den Brom R, Dercksen D (2010): Research into possible side effects of Coxevac. Deventer.
Verhasselt V, Vosters O, Beuneu C, Nicaise C, Stordeur P, Goldman M (2004): Induction of FOXP3-expressing regulatory CD4pos T cells by human mature autologous dendritic cells. Eur J Immunol 34: 762–772.
Vila-del Sol V, Diaz-Munoz MD, Fresno M (2007): Requirement of tumor necrosis factor alpha and nuclear factor-kappaB in the induction by IFN-gamma of inducible nitric oxide synthase in macrophages. J Leukoc Biol 81: 272–283.
von Korn S (2001): Schafe in Koppel- und Hütehaltung. Eugen Ulmer Verlag, Stuttgart.
Voth DE, Heinzen RA (2007): Lounging in a lysosome: the intracellular lifestyle of Coxiella burnetii. Cell Microbiol 9: 829–840.
Voth DE, Howe D, Heinzen RA (2007): Coxiella burnetii inhibits apoptosis in human THP-1 cells and monkey primary alveolar macrophages. Infect Immun 75: 4263–4271.
Walter M, Brugger K, Rubel F (2016): The ecological niche of Dermacentor marginatus in Germany. Parasitol Res 115: 2165–2174.
Weber MM, Faris R, van Schaik EJ, McLachlan JT, Wright WU, Tellez A, Roman VA, Rowin K, Case EDR, Luo Z-Q, Samuel JE (2016): The type IV secretion system effector protein CirA atimulates the GTPase activity of RhoA and is required for virulence in a mouse model of Coxiella burnetii infection. Infect Immun 84: 2524–2533.
Wegdam-Blans M, Kampschreur L, Delsing C, Bleeker-Rovers C, Sprong T, Van Kasteren M, Notermans DW, Renders N, Bijlmer H, Lestrade P (2012): Chronic Q fever: review of the literature and a proposal of new diagnostic criteria. J Infection 64: 247–259.
Weisburg WG, Dobson ME, Samuel JE, Dasch GA, Mallavia LP, Baca O, Mandelco L, Sechrest JE, Weiss E, Woese CR (1989): Phylogenetic diversity of the Rickettsiae. J Bacteriol 171: 4202–4206.
Welsh HH, Lennette EH, Abinanti FR, Winn JF (1951): Q fever in California: IV. Occurrence of Coxiella burnetii in the placenta of naturally infected sheep. Public Health Rep 66: 1473–1477.
Winchell CG, Graham JG, Kurten RC, Voth DE (2014): Coxiella burnetii type IV secretion-dependent recruitment of macrophage autophagosomes. Infect Immun 82: 2229–2238.
Woldehiwet Z (2004): Q fever (coxiellosis): epidemiology and pathogenesis. Res Vet Sci 77: 93–100.
Zamboni DS, Campos MA, Torrecilhas AC, Kiss K, Samuel JE, Golenbock DT, Lauw FN, Roy CR, Almeida IC, Gazzinelli RT (2004): Stimulation of toll-like receptor 2 by Coxiella burnetii is required for macrophage production of pro-inflammatory cytokines and resistance to infection. J Biol Chem 279: 54405–54415.
Zeman DH, Kirkbride CA, Leslie-Steen P, Duimstra JR (1989): Ovine abortion due to Coxiella burnetii infection. J Vet Diagn Invest 1: 178–180.
Zhang Y, Zhang G, Hendrix LR, Tesh VL, Samuel JE (2012): Coxiella burnetii induces apoptosis during early stage infection via a caspase-independent pathway in human monocytic THP-1 cells. PLoS One 7: e30841.
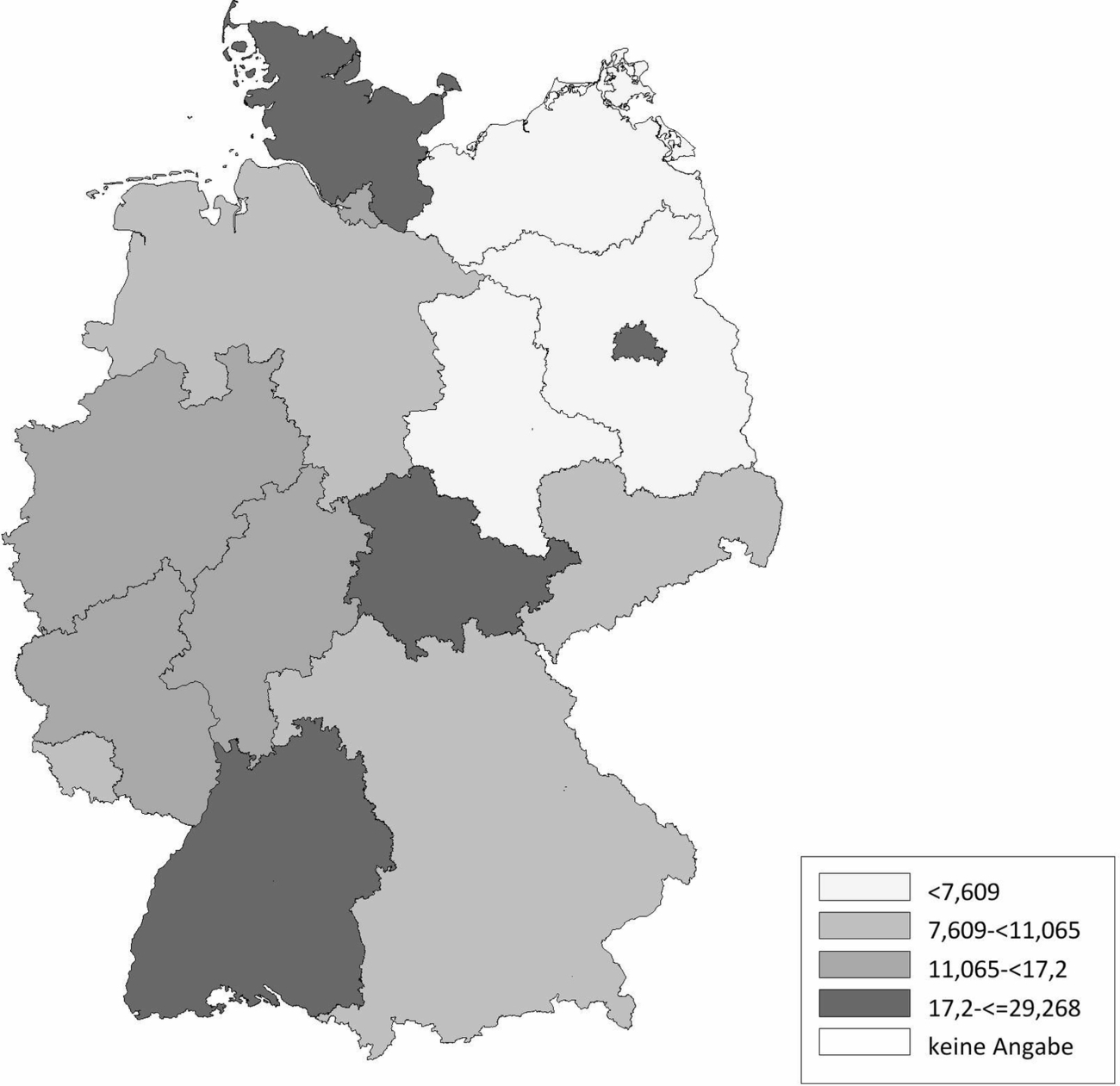
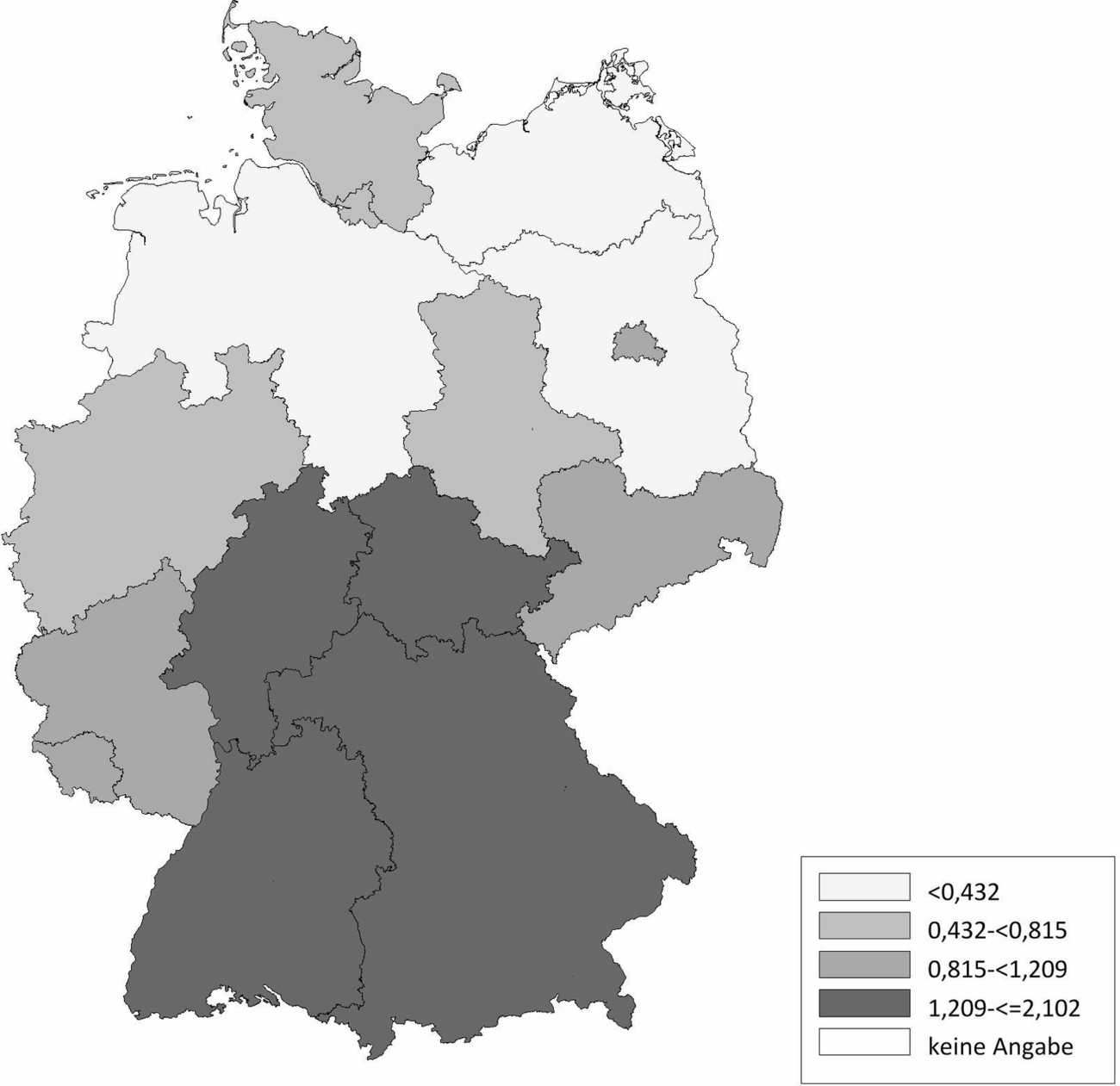
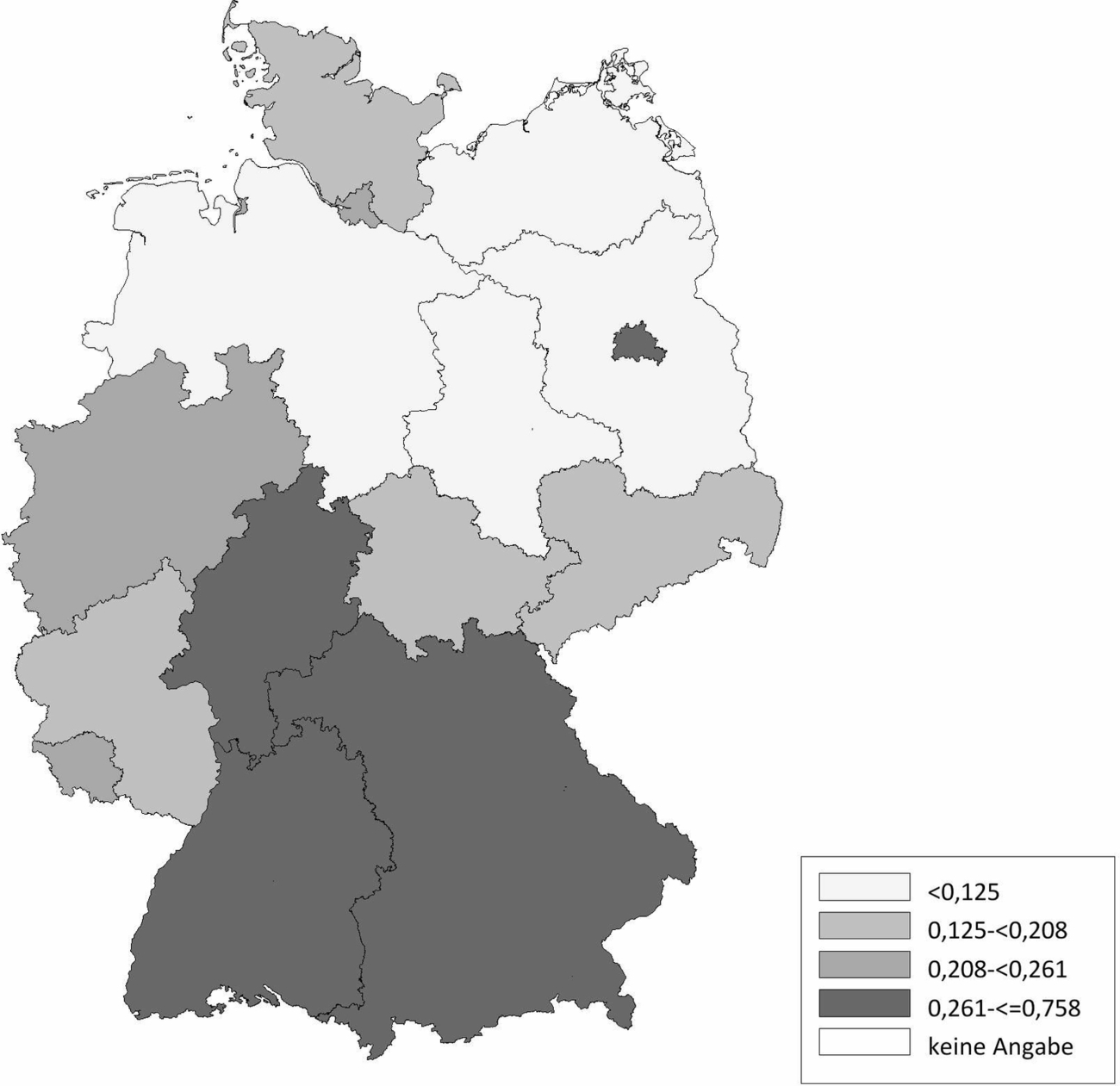
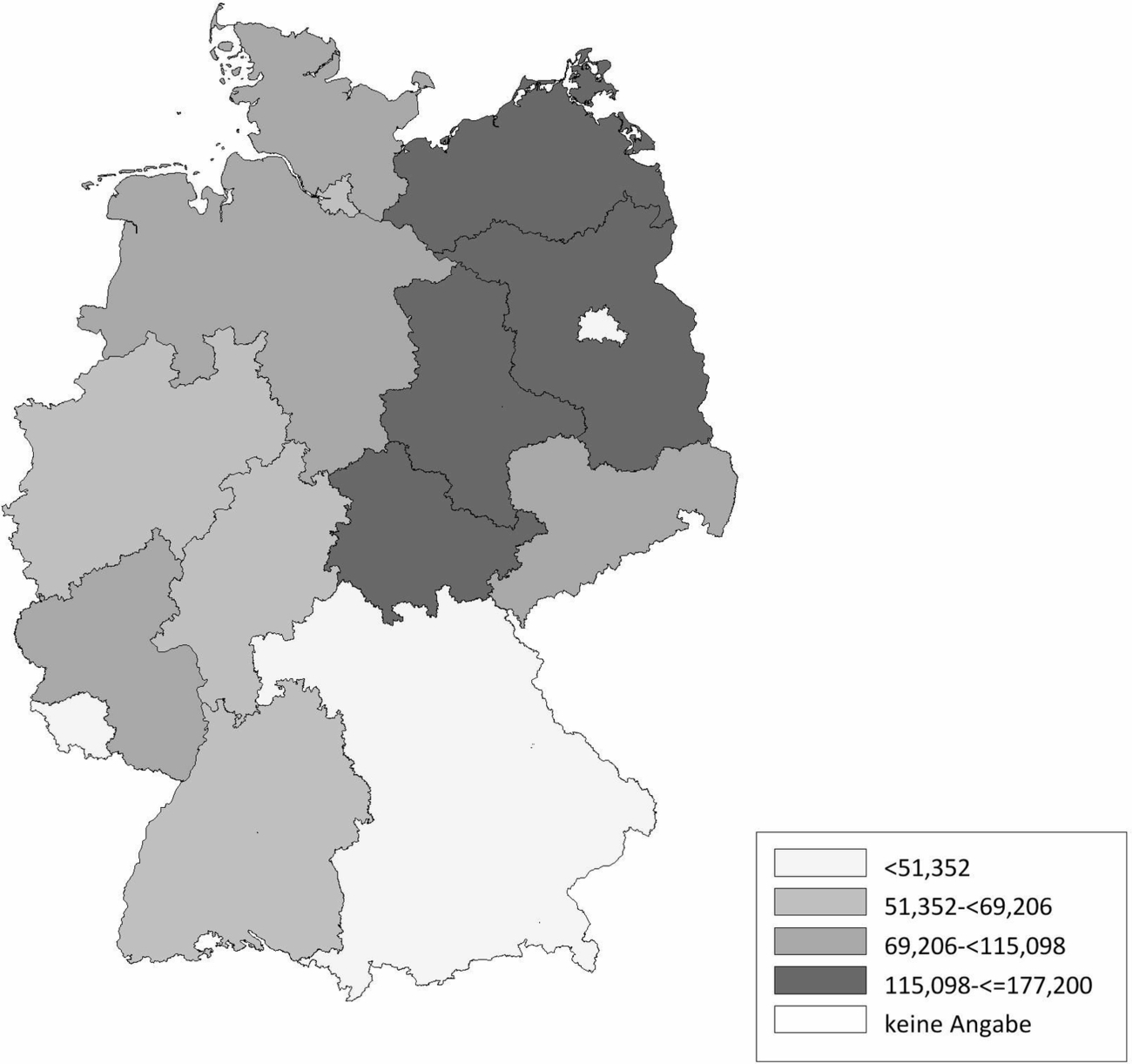
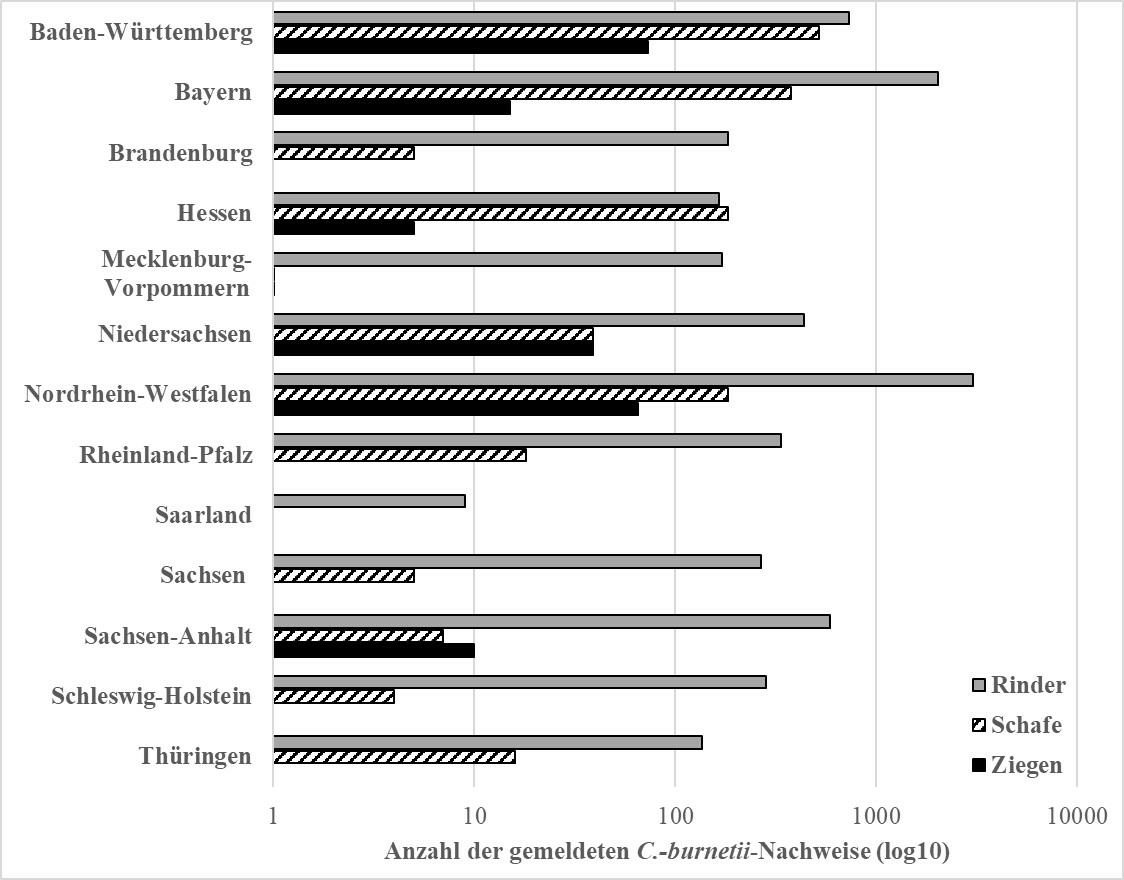


Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23760005-9366-19030-Bauer.pdf (0.47 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23760005-9366-19030-Bauer-Tabelle 1.pdf (0.13 MB) herunterladen möchten


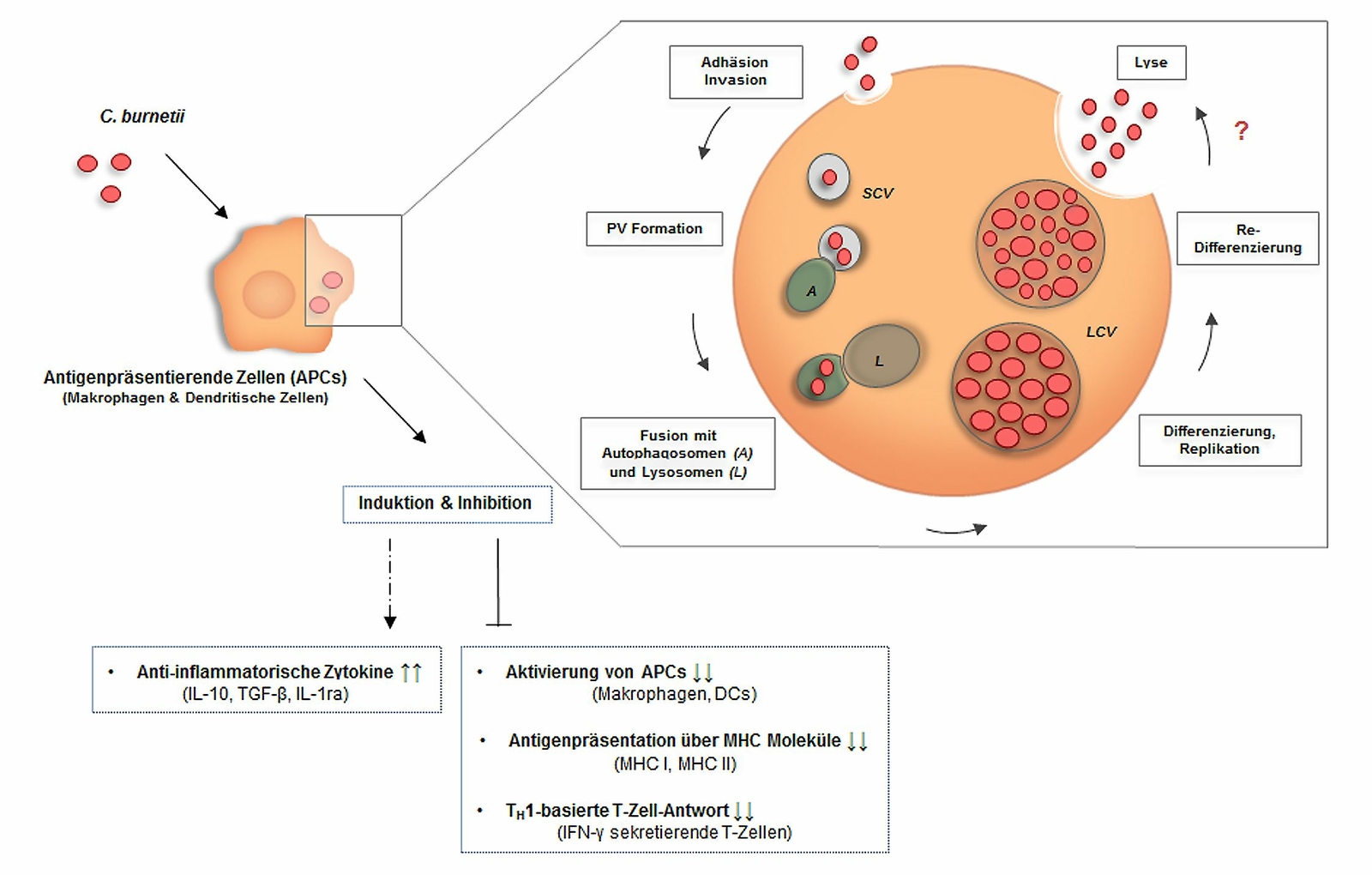{kind=link}
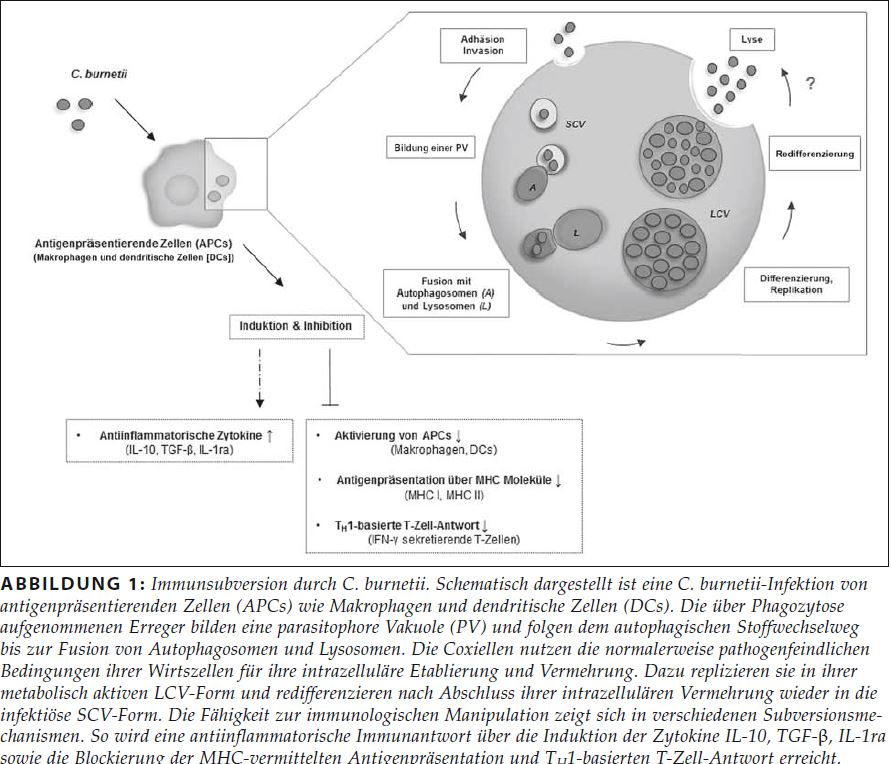{kind=link}
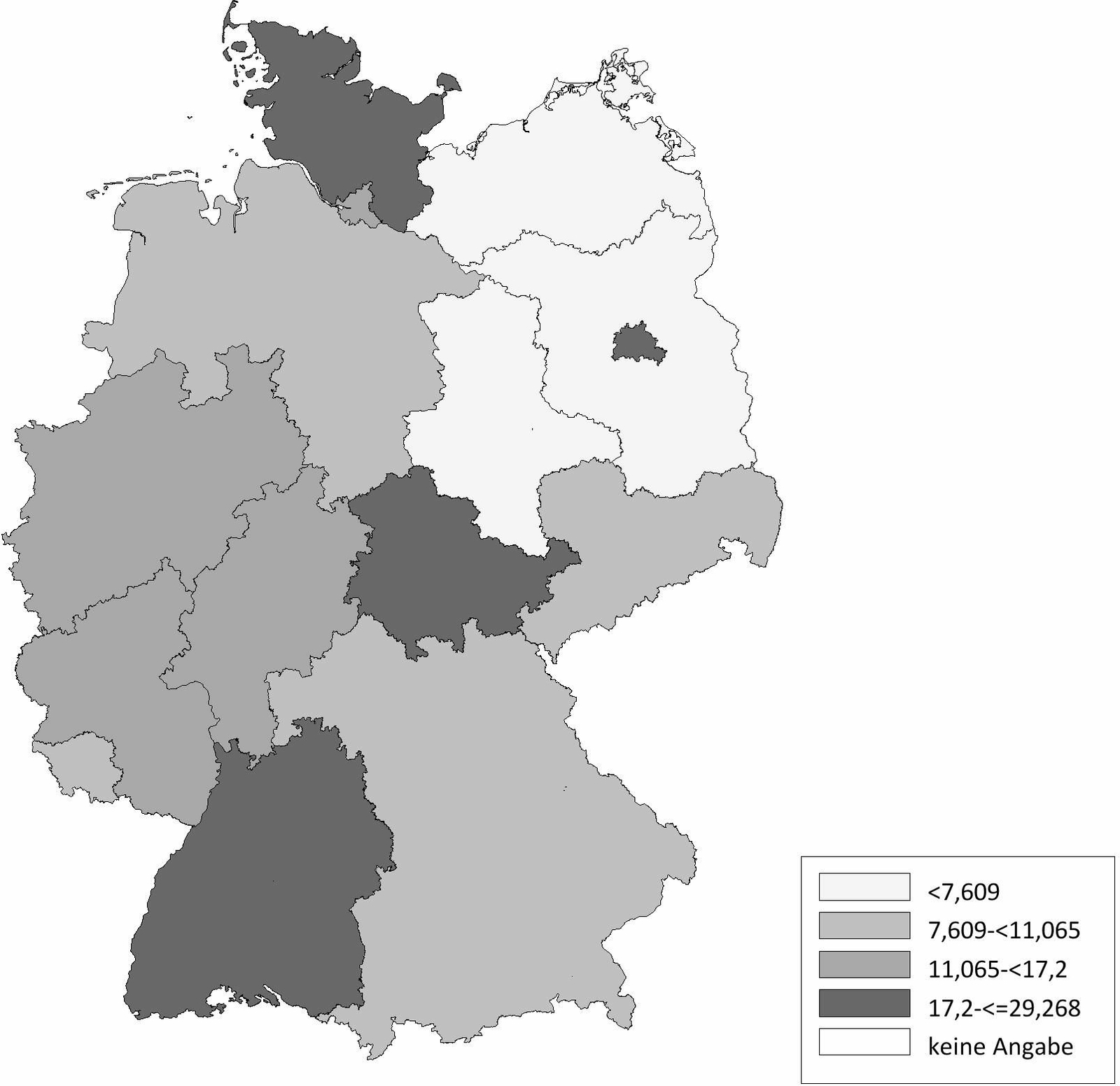{kind=link}
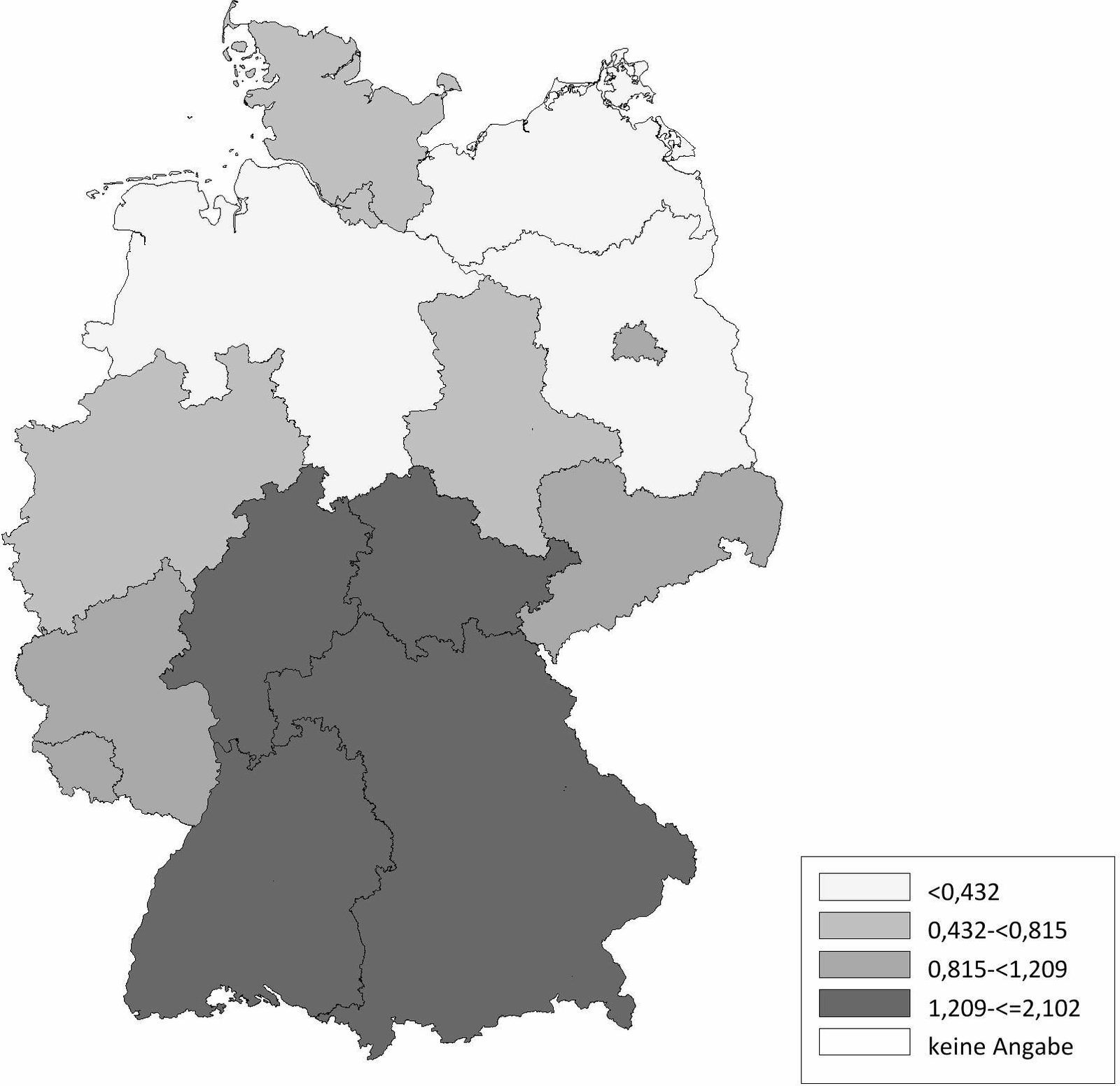{kind=link}
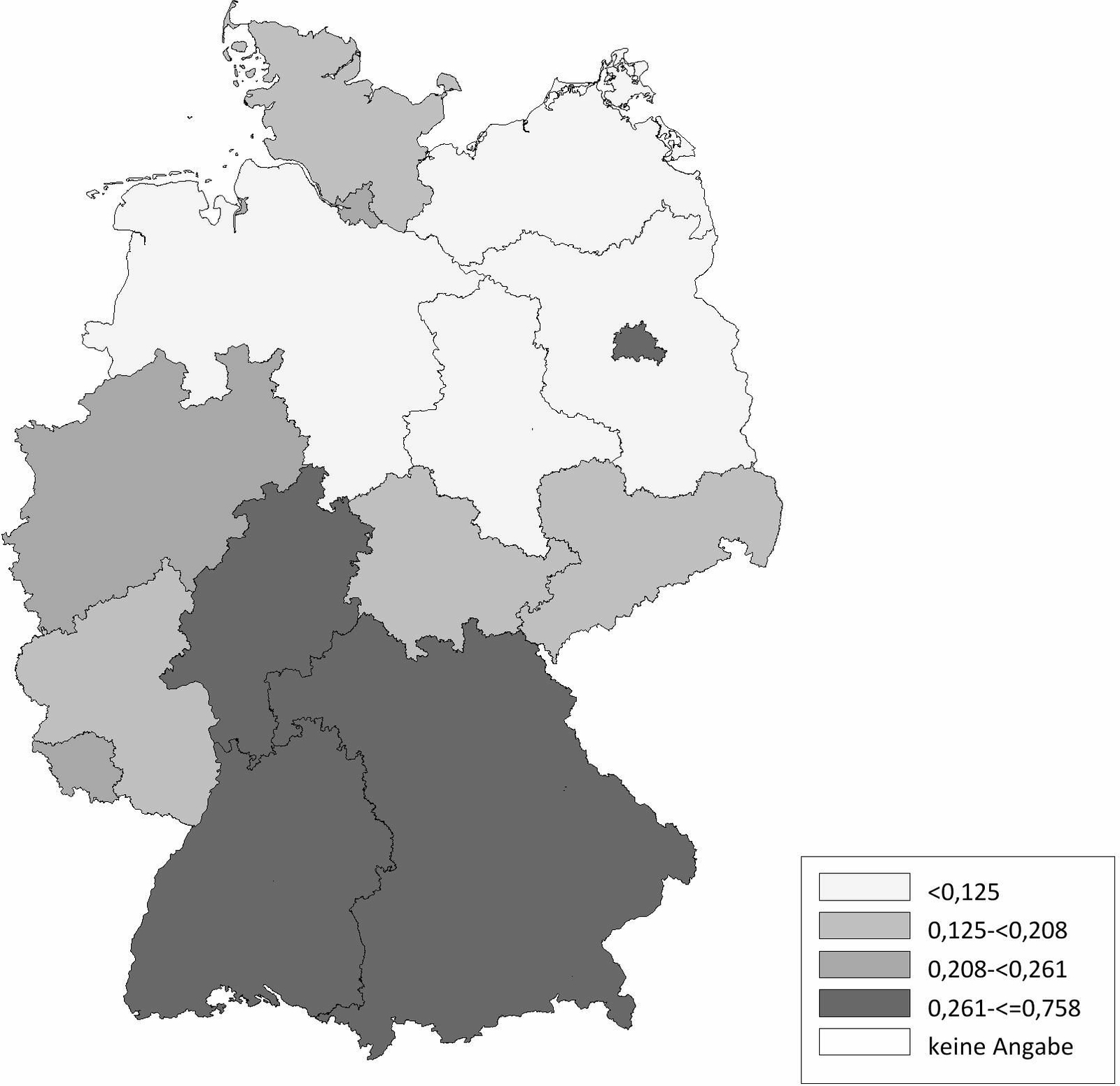{kind=link}
{kind=link}
{kind=link}
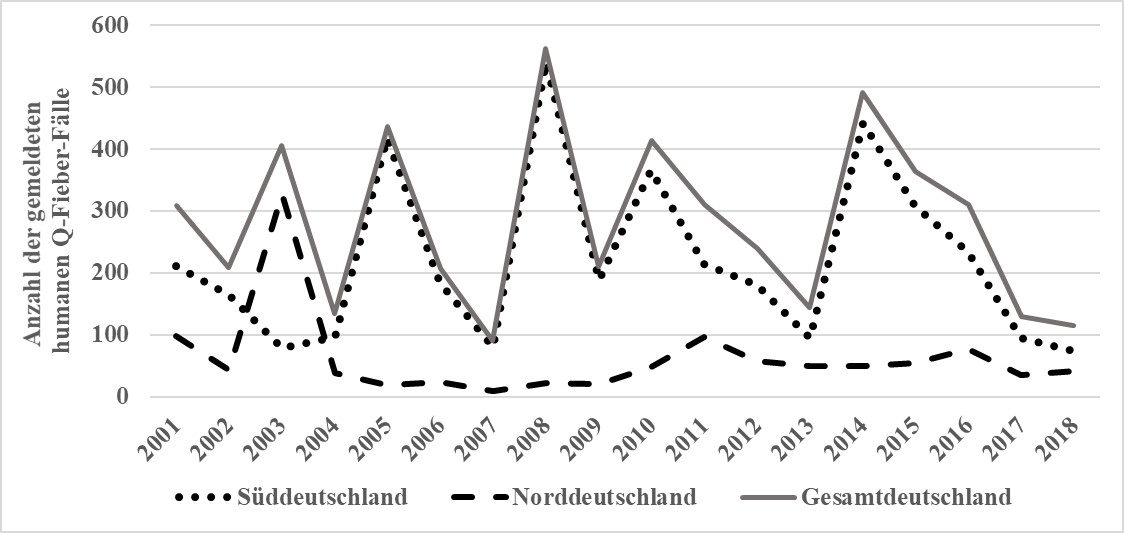{kind=link}