

Nachweis einer Mycobacterium avium subsp. hominissuis INMV 51-Infektion bei einem Rotschopfturako (Tauraco erythrolophus)
Berliner und Münchener Tierärztliche Wochenschrift 134, 1-7
DOI: 10.2376/1439-0299-2020-39
© Schlütersche Fachmedien GmbH. 2021
Publiziert: 02/2021
Summary
A four years old Red-crested turaco (Tauraco erythrolophus) was found apathetic sitting on the ground of the aviary and died during the clinical examination. Necropsy of the emaciated bird revealed multiple granulomas within the body cavity and inner organs. Histopathological examinations of these granulomas showed epitheloid macrophages and central necrosis demarcated by giant cells. Using Ziehl-Neelsen staining, massive accumulations of acid-fast bacteria were found within the lesions and the preliminary diagnosis “avian tuberculosis” was made. This infectious disease is mainly caused by Mycobacterium avium subsp. avium (Maa) or M. genavense (Mg). However, microbiological culture followed by PCR revealed the occurrence of Mycobacterium avium subsp. hominissuis (Mah). The isolate was assigned to INMV profile 51, which has been described in Asia and Europe mainly in humans, as well as in pigs and cattle until now. When infected birds have intensive contact with humans, a risk assessment must be carried out based on the identity of the causative pathogen. For this purpose, the determination of the species and if applicable the subspecies is mandatory and an additional molecular typing, e.g. by INMV profiling, helpful.
Zusammenfassung
Ein vier Jahre alter Rotschopfturako (syn. Rothaubenturako, Tauraco erythrolophus) wurde apathisch auf dem Boden der Voliere sitzend aufgefunden und verstarb während der klinischen Untersuchung. Bei der Sektion des abgemagerten Vogels wurden multiple Granulome in der Körperhöhle und den inneren Organen festgestellt. Histopathologische Untersuchungen der Granulome zeigten epitheloide Makrophagen und zentrale Nekrosen, die durch Riesenzellen abgegrenzt waren. Mittels Ziehl-Neelsen-Färbung wurden massive Anhäufungen von säurefesten Bakterien in den Läsionen gefunden und die vorläufige Diagnose „Geflügeltuberkulose“ gestellt, eine Infektionskrankheit, die hauptsächlich durch Mycobacterium avium subsp. avium (Maa) oder M. genavense (Mg) verursacht wird. Die mikrobiologische Kultur und PCR ergaben im vorliegenden Fall aber das atypische Vorkommen von Mycobacterium avium subsp. hominissuis (Mah) bei dem Turako. Das Isolat wurde dem INMV-Profil 51 zugeordnet, das bisher vor allem in Asien und Europa beim Menschen, aber auch bei Schweinen und Rindern beschrieben wurde. Wenn infizierte Vögel intensiven Kontakt zu Menschen haben, muss eine Risikobewertung auf der Grundlage der Identität des ursächlichen Pathogens durchgeführt werden. Hierfür sind die Bestimmung der Spezies und ggf. Subspezies obligatorisch und eine molekulare Feintypisierung, z. B. mittels INMV-Typisierung, hilfreich.
Introduction
Red-crested turacos are middle-sized birds of the family Musophagidae. They are endemic to Angola’s forests where these territorial, frugivorous birds live in trees. They occur in smaller groups or pairs and live mostly monogamously. Red-crested turacos can reach an age of five to nine years in the wild and in captivity up to 30 years (Tocidlowski 2012). Because of their colourful plumage, turacos are often kept in zoos and enjoy great popularity (Peat 2017). According to the IUCN Red List of Threatened Species the wildlife population density is decreasing, but the status is still “least concern” (BirdLife International 2016).
Mycobacteria are aerobic growing rod-shaped bacteria, which are characterized by a waxy mycolic acid lipid layer in the outer cell membrane that mediates the acid-fast phenotype in different staining methods (e.g., Ziehl-Neelsen, Kinyoun, Fite-Faraco). Compared to other bacteria, most Mycobacterium (M.) species grow very slowly in vitro, so that cultural detection can take several months. Only very few members of the genus are obligate pathogens for humans, in particular the members of the M. tuberculosis complex (MTC) and some non-tuberculous mycobacteria (NTM), e.g. M. ulcerans or M. leprae, while most NTM can be found ubiquitous in the environment, e.g., in soil and water, and are facultative pathogenic for their respective hosts.
In captive birds and poultry avian tuberculosis (avTB) is a chronic infectious disease that occurs worldwide (Dvorska et al. 2007). The clinical signs remain unspecific in most cases and may include emaciation, apathy, lameness, ruffled feathers and/or diarrhoea. Nevertheless, also cases without any premortal clinical signs are described (Tell et al. 2001). Contrary to what the name “tuberculosis” suggests, the disease is caused by NTM. In poultry avTB is mainly caused by M. avium subsp. avium (Maa), whereas pet birds are mostly infected by M. genavense (Mg) (Manarolla et al. 2009). Beside the previous mentioned two major pathogens of avTB, other NTMs (e.g., M. fortuitum, M. intracellulare, M. scrofulaceum, M. gordonae, M. nonchromogenicum, M. celatum, M. intermedium) can cause identical pathological lesions in affected birds (Bertelsen et al. 2006, Hoop et al. 1996, Kik et al. 2010, Pfeiffer et al. 2017). Rarely infections with members of the MTC, e.g. M. tuberculosis or M. bovis, are reported in birds (Hoop et al. 1996, Peters et al. 2007, Sanchez et al. 2016, Steinmetz et al. 2006).
Top Job:
In the early ‘90s Mg was first detected in immunodeficient humans, especially AIDS patients, and in pet birds (Böttger et al. 1993, Hoop et al. 1993). The culture of Mg is difficult, as it demands high standards to the media (supplementation of mycobactin J), the incubation atmosphere (5% CO2, 42°C) and needs long incubation periods (up to several months) (Böttger et al. 1993). Therefore, molecular methods like polymerase chain reaction (PCR) are of high importance in diagnostic workflows. Several protocols are published (Chevrier et al. 1999, Mendenhall et al. 2000, Tell et al. 2003). Mainly pet birds, such as amazon parrots, zebra finches, budgerigars or other psittacids (Ferrer et al. 1997, Gomez et al. 2011, Hoop et al. 1993), but also free-living birds may be infected by Mg (Patino et al. 2018). Besides various avian species and humans, Mg was found in several other mammalians, e.g. horses, ferrets, squirrels or chinchillas, or even in reptiles, e.g. snakes (Bottcher et al. 2016, De Lorenzi et al. 2018, Huynh et al. 2014, Theuss et al. 2010, Ullmann et al. 2016).
According to genetic examinations, M. avium subsp. bacteria belong to a complex comprising 12 species (van Ingen et al. 2018). Nevertheless, in the strict sense the M. avium complex (MAC) focussed in the past on the four M. avium subspecies avium, silvaticum (Mas), hominissuis (Mah) and paratuberculosis (Map), as well as M. intracellulare (Mi) (Biet et al. 2005, Turenne et al. 2007). Thereof, Maa and Mas are mainly found in avians, Mah and Mi in a variety of host species and environmental samples, whereas Map causes the so-called Johne’s disease in ruminants (Turenne et al. 2007). The MAC members can be differentiated by their dependence of the growth factor mycobactin J, mandatory for in vitro growth of Map and Mas, and the serological reactivity in serovars 1-3 (Maa), serovars 4-6, 8-11 and 21 (Mah) and serovars 7, 12-20 and 22-28 (Mi) (Wayne et al. 1993). Today, the presence or absence of specific insertion sequences (IS) is tested by different PCRs and allows a much faster assignment to the respective M. avium subspecies: IS1311 (present in all M. avium subsp.), IS1245 (present in Maa, Mas, Mah), IS901/IS902 (present in Maa, Mas) and IS900 (present in Map) (Turenne et al. 2007).
Single cases of mycobacterial infections are described in turacos from zoological gardens in the U.S.A., with only unspecific clinical signs such as poor body condition or soft subcutaneous swellings in the birds (Brannian 1993, Stamper et al. 1998, Tocidlowski 2012, Wilson and Carpenter 1994). Thereby, the frequency with which mycobacterial infections are claimed to be responsible for the death of individuals in the turaco populations varied between the zoos from 3% to 66%, indicating that turacos are not particularly susceptible to mycobacteria (Stamper et al. 1998, Tocidlowski 2012). The diagnosis avian tuberculosis was mainly based on gross and histopathological findings and only in individual cases confirmed by mycobacterial culture, subsequently resulting in the species diagnosis MAC. In the remaining cases, a mycobacterial species more difficult to cultivate, e.g. Mg, may have been the causative agent, likewise it was shown in one Great Go-Away-Bird, also a member of the family Musophagidae (Pfeiffer et al. 2017, Stamper et al. 1998, Wilson and Carpenter 1994).
The current case report describes the occurrence of avian tuberculosis in a Red-crested turaco, atypically caused by the mycobacterial subspecies Mah.
Case description
Animal
A four years old, female, Red-crested turaco (Tauraco erythrolophus) was found apathetic sitting on the ground of its enclosure and died during the clinical examination. The bird hatched in 2016 at a zoological institution in Great Britain, which was approved according to the Council Directive 92/65/EEC for the trade of animals within the EU, and imported at the end of the year 2018 into the Berlin Zoological Garden. There it was housed in a subunit of an aviary called the “Fasanerie” together with its male partner, 30 Java sparrows (Lonchura oryzivora), two Harlequin quails (Coturnix delegorguei), two Golden-breasted starlings (Lamprotornis regius), two Spotted thick-knees (Burhinus capensis), and one Malagasy turtle dove (Nesoenas picturata). The birds are kept under conditions allowing inside/outside access without contact to visitors. They have free access to food prepared by the zookeepers consisting of apple, pear, orange, grapes, cherries, cooked corn, tomato, cucumber, bell pepper, mealworms and bird minerals (Salvana Tiernahrung, Kl. O.-Sparrieshoop) and water. The aviary has a natural ground allowing exposure to the soil of the enclosure. Direct contact with birds from the other subunits of the “Fasanerie” or other aviaries of the zoo is impossible. However, indirect contact via the zookeepers or equipment used for maintenance of the enclosures can´t be ruled out. In addition, contact with the excretions of wild birds and rodents passing through the mesh fence of the subunits may happen. All living birds are currently without clinical signs. Birds of various other avian species are generally kept in caged aviaries in the zoo (Table 1), except for Sarus cranes (Antigone antigone), which have access to a fenced green land enclosure, allowing contact with wild birds and mammals. At the Berlin Zoological Garden, avTB caused by Maa and Mg but not by Mah has repeatedly been diagnosed in various subunits of the “Fasanerie” and the other aviaries in the past (Table 1). However, the turaco represents the first case of avTB in its subunit and the first case of Mah associated avTB in this zoo.
Pathologic-anatomic and histologic examination
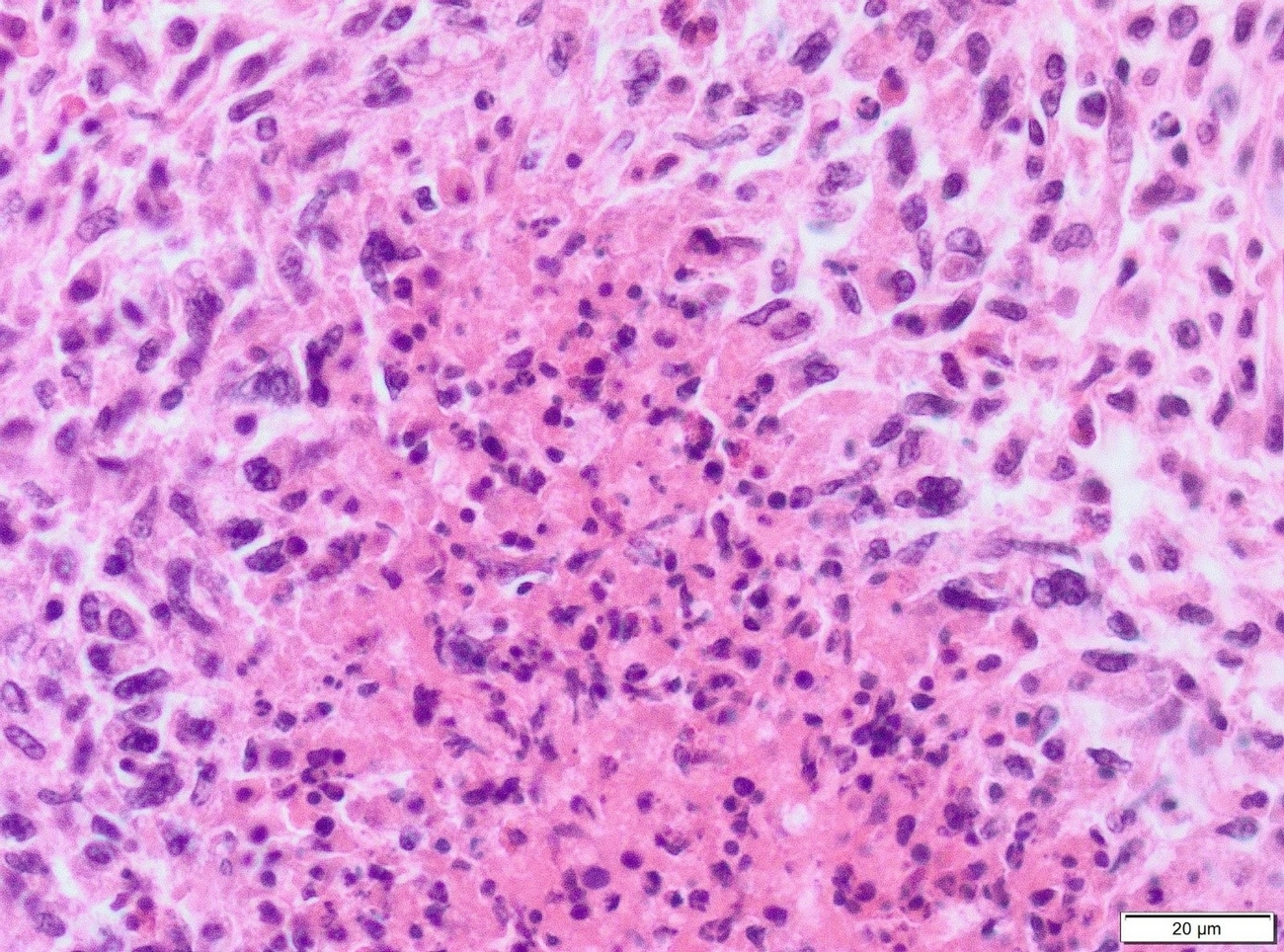
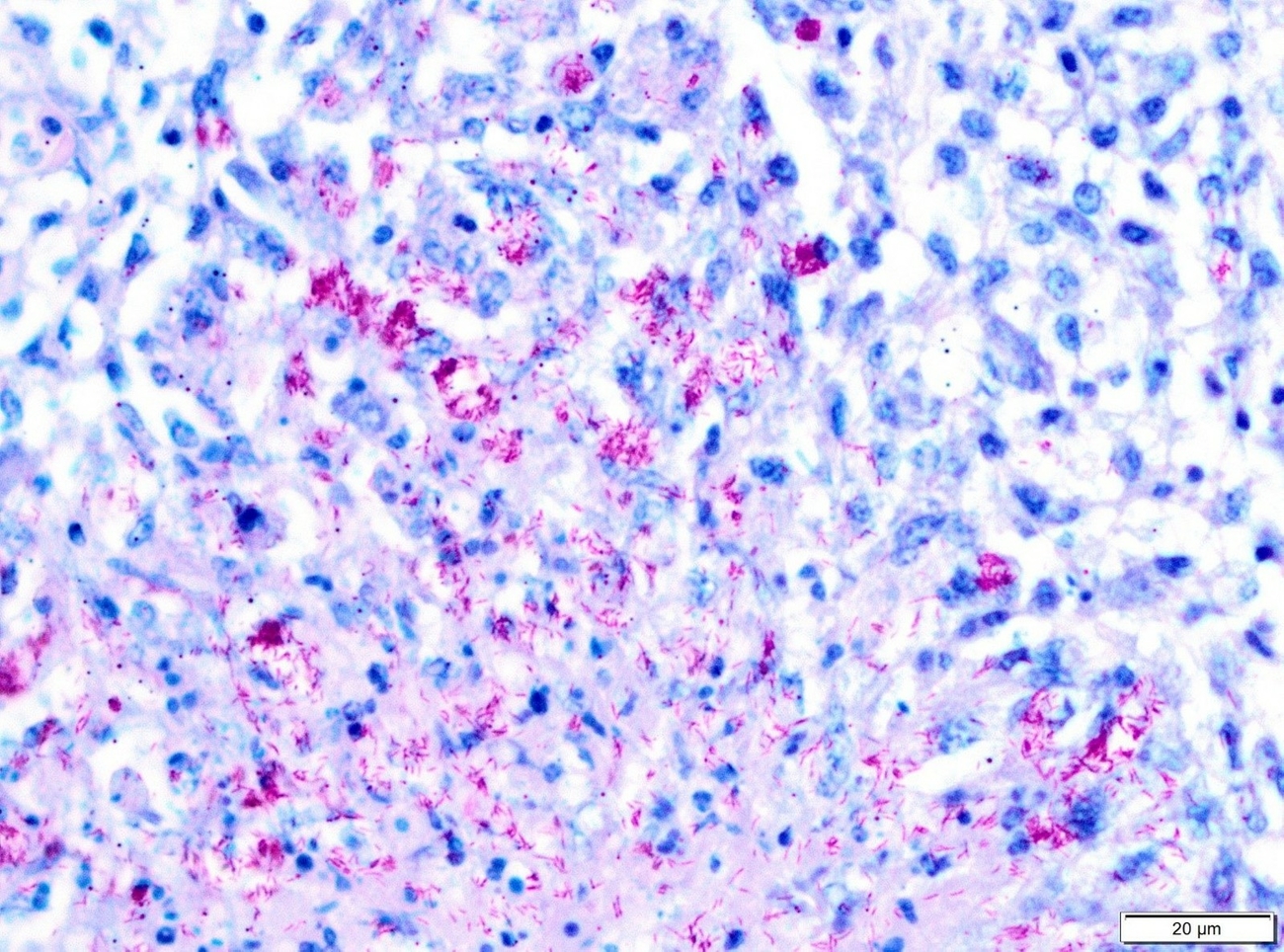
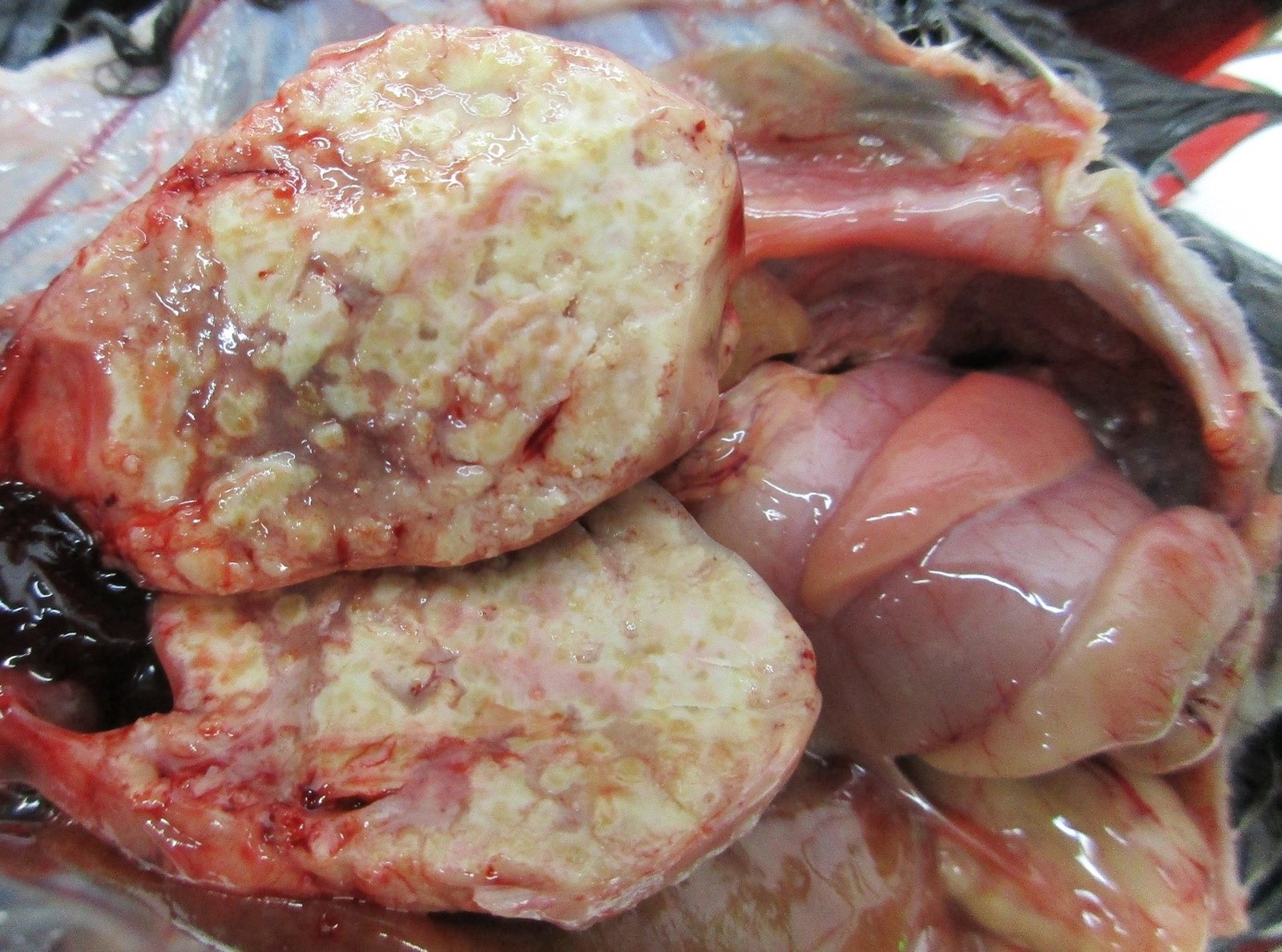
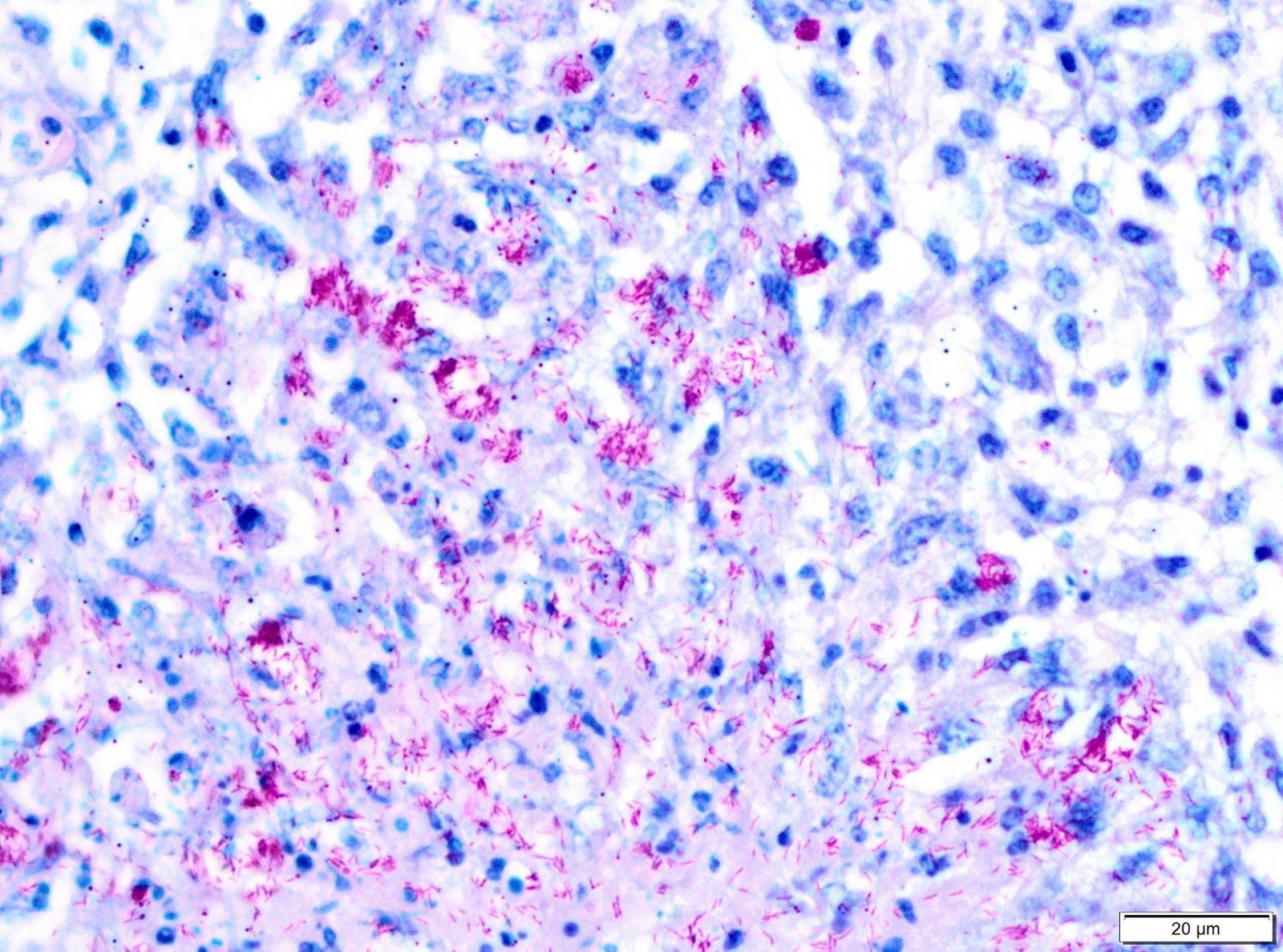
Necropsy of the emaciated bird revealed multiple granulomas of up to 2 cm in diameter within the body cavity (Figure 1). In addition, miliary confluating granulomas were present in various inner organs especially spleen, liver and lung with destruction of approximately 80% of the lung parenchyma. Massive amounts of acid-fast bacteria were detected in Ziehl-Neelsen-stained imprint smear preparations of the granulomas. Histopathologically, the lesions were characterized by an infiltration of epitheloid macrophages and the formation of granulomas with central necrosis demarcated by giant cells (Figure 2a). Again, acid-fast bacteria were found in the cytoplasm of the epitheloid macrophages, as well as extracellularly within the necrotic areas by Ziehl-Neelsen staining (Figure 2b). Granulomas were detected neither macroscopically within the whole digestive tract nor in routine histopathological specimens from proventriculus, ventriculus and jejunum.
Microbiological examinations
Concurrent bacterial, mycotic and parasitic infections were ruled out by routine diagnostic workup including bacteriology of heart, lung, liver and intestine using standard culture media, mycology of the lung and coproscopical examination of intestinal contents for the detection of developmental stages of endoparasites. In addition, infections with chlamydiae, Influenza A virus, Avian orthoavulavirus 1, West Nile virus and Usutu virus were ruled out by PCR and virus culture using standard protocols approved by the German veterinarian National Reference Laboratories (Friedrich-Loeffler-Institut 2019).
Identification and characterization of the mycobacterial isolate
Ziehl-Neelsen-staining of putative mycobacterial bacterial culture grown on Stonebrink agar slants showed the presence of acid-fast rods. From the colony material DNA was extracted using a heat and ultra-sonic lysis of the bacterial cells. For species and subspecies identification the DNA was submitted to PCR analysis targeting Mycobacterium sp.-specific 16S rRNA, M. avium subsp.-specific insertion sequences (IS), which included IS1245, IS900, and IS901 (Friedrich-Loeffler-Institut 2019), as well as a Mg-specific DNA fragment (Chevrier et al. 1999). Thereby, the isolate was identified to be Mah, as it was positive for mycobacterial 16S rRNA and IS1245, but negative for IS901, IS900 and the Mg-specific DNA fragment.
The Mah isolate was further characterized by genotyping using the Mycobacterial Interspersed Repetitive Units-Variable Number Tandem Repeat (MIRU-VNTR) analysis, encompassing the loci VNTR 292, X3, 25, 47, 3, 7, 10, and 32, following the protocol of Thibault et al. (2007) and the MAC-INMV-SSR database (http://mac-inmv.tours.inra.fr) (Cochard et al. 2020). The identified pattern of repetitive counts (2-4-2-2-1-1-2-8) matched the profile INMV 51.
Discussion
In captive birds and poultry avTB is a chronic infectious disease that occurs worldwide (Dvorska et al. 2007). In commercial holdings the disease is of decreasing importance, but in the last years poultry in smaller backyard farms and pet birds in private households seem to be affected more often (Tell et al. 2001). In addition, avTB is a long known problem in various avian species from zoological holdings (Keymer et al. 1982, Krause et al. 2015, Montali et al. 1976, Portaels et al. 1996, Witte et al. 2008). Ante mortem diagnosis in birds, based on tuberculin tests, serology, haematology or radiography, is often not conclusive to identify individual animals and, especially the immunological assays, more suitable at herd level (Montali et al. 1976, O.I.E. 2018). The tuberculin test, performed by application of purified protein derivate (PPD) of Maa in the skin of the wattle, comb or the propatagium, and assessed 48 h later for inflammation, is unreliable in several avian species and mainly used for pre-movement testing of domestic fowl (O.I.E. 2018, Tell et al. 2001). Consequently, diagnosis of avTB in individuals is often based on the pathological findings during necropsies and detection of the pathogen by culture and/or molecular diagnostics, e.g., polymerase chain reaction (PCR). Even though the pathological findings can be very diverse, the presence of granulomas or accumulations of epitheloid macrophages in single or multiple organs in combination with acid-fast rods within the lesions are pathognomonic. Most often multiple organs, such as liver, lung, spleen and/or intestine are affected in birds suffering from avTB, as in the case described here (Hoop et al. 1993, Tell et al. 2001, Witte et al. 2008). However, disseminated forms without visible granulomas at necropsy or cases with affection of only single organs (e.g., eye-lids, skin or bones) are known (Montali et al. 1976, Portaels et al. 1996).
Data sets from zoos, where dead birds are often routinely necropsied, show that repeated cases of avTB can occur in a certain flock, but often only single individuals are affected (Portaels et al. 1996, Witte et al. 2008). At the Berlin Zoo, 15 animals with avTB, in which the mycobacterial species was identified, were diagnosed from 2014 onwards, affecting five different aviaries. Overall, in six birds Mg was the causative agent, in eight birds Maa, and in the case described here Mah. Besides a study of Pfeiffer et al. (2017) at the San Diego Zoo that identified Mah in 11.4 % of 105 birds with avTB by whole genome sequencing, only single descriptions of Mah infections in birds exist, e.g. in a blue-fronted Amazon parrot (Shitaye et al. 2009) or in a painted quail (Morita et al. 1999). The Mah isolates from the quail were subsequently used to infect chickens experimentally with 3-5 x 107 cfu/dose (Morita et al. 1999). The infection resulted in dramatic weight-loss and lethargy in all immunocompetent chicken within 44 days, proving the pathogenicity of Mah for birds.
The MIRU-VNTR analysis of the isolate from the turaco assigned it to profile INMV 51. This profile was until now only described in isolates from humans, cattle and pigs in Europe and Asia (Iwamoto et al. 2012, Radomski et al. 2010, Scherrer et al. 2018, Starkova et al. 2013). In fact, INMV 51 was the predominant profile within the human isolates tested in a French study (Thibault et al. 2007), demonstrating the zoonotic potential of the Mah type detected in the turaco representing the first report of INMV 51 in an avian host. The presence of INMV 51 in several studies on different continents might be a hint for a broad distribution of this Mah type in the environment or perhaps in wild birds, which may act as transport vehicles over long distances.
The origins of infection in most cases of avTB are usually not known, but the environment and/or other animals are the most likely sources. Members of the MAC are known to survive for long time periods in the environment, especially in soil and water (Lahiri et al. 2014). Furthermore, the environment and tap water are suspected reservoirs of Mg (Portaels et al. 1996). The aviaries in the Berlin Zoo have soil floors to promote the natural behaviour of the birds, and all affected holdings contain inside and outside areas, where contact with droppings of wild birds or rodents is possible. This means that by keeping the birds under near natural conditions, which is urged by reasons of animal welfare, they have a risk to come in contact with environmental pathogens. Maa, Mah and Mg have been detected in faecal samples or gastrointestinal tract of several bird species (Bercovier and Vincent 2001, Morita et al. 1999, Patino et al. 2018, Shitaye et al. 2009, 2010, Smit et al. 1987) and rodents (Maa) (Fischer et al. 2000). Therefore, the faecal excretion of NTM is an important source of infection for other birds, animals or humans directly or indirectly through contamination of the environment, food or water. Already Witte et al. (2008) showed that birds living together with individuals suffering from intestinal mycobacteriosis have a higher infection risk. More specifically, Mah was shown to be present in the intestine of a painted quail and a blue-fronted Amazon parrot as well as in the feces of experimentally infected chickens (Morita et al. 1999, Shitaye et al. 2009) and the introduction of new birds into an existing flock was a risk factor to develop avTB in zoo birds (Witte et al. 2008). In this context, comparable to Trojan horses, seemingly healthy birds can be mycobacterial carriers and may develop clinical disease long time after their import or infect other birds in the flock. In addition, stress that occurs when new animals are introduced into an existing flock can trigger an immunosuppression and an already present pathogen can cause clinical disease. Since Mah is a nearly ubiquitous pathogen that potential hosts have constantly to deal with, a lowered immune response is often a prerequisite for a clinically apparent infection. Summarizing, the exacerbation of a subclinical infection with Mah was considered to be the most likely explanation for the case of Mah related avTB presented here, although, the definitive source of the infection cannot be determined anymore.
An interesting observation in the zoo is that Mg infections were only detected in birds of the “Afrikaloop” of the bird house “World of Birds”, while Maa infections were found in all locations with cases of avTB. The concentration of Mg infections in only one aviary can be interpreted as an indication of an animal-to-animal transmission. This hypothesis is supported by data from whole genome analyses of Mg and Maa isolates from birds within San Diego Zoo (Pfeiffer et al. 2017), where 48 Mg isolates showed a significant lower genome variability among each other than the parallel tested Mah (n=12) and Maa (n=37) isolates possessed even on subspecies level. Whereas, the genetic heterogeneity of the MAC isolates suggested a repeated entry of the pathogens into the population and, accordingly, less frequent animal-to-animal transmission (Pfeiffer et al. 2017, Schrenzel et al. 2008).
Finally, avTB can be caused by different mycobacterial species and subspecies. If infected birds have intense contact to their owner or in zoological gardens with animal care takers or even visitors, a hazard assessment should be carried out based on the identification and typing of the causing pathogen. While Maa infections in humans are rare, Mah and Mg can be isolated from immunosuppressed and -competent humans (Cornelis et al. 2018, Martin and Schimmel 2000, Möbius et al. 2006, Tran and Han 2014). Accordingly, the zoonotic risk may be considered higher for Mah or Mg than for Maa.
Ethical approval
The authors assure that they have followed the general rules of good scientific practice.
Conflict of interest
The authors assure that there are no proprietary, professional or other personal interests in any product or company that could influence the content or opinions expressed in this publication.
Authors contribution
Concept: SAB, CS.
Data collection: AO, CS, NW, SB, SAB, PM.
Analysis and interpretation: AO, CS, SB, NW, SAB, PM.
Draft manuscript: SAB.
Critical revision: CS, PM, AO.
Final approval of the version intended for publication: AO, CS, SB, NW, SAB, PM.
Address for correspondence
Dr. Stefanie A. Barth
Friedrich-Loeffler-Institut/Bundesforschungsinstitut für Tiergesundheit
Institut für molekulare Pathogenese
Naumburger Str. 96A
07743 Jena, Germany
stefanie.barth@fli.de
References
Bercovier H, Vincent V (2001): Mycobacterial infections in domestic and wild animals due to Mycobacterium marinum, M. fortuitum, M. chelonae, M. porcinum, M. farcinogenes, M. smegmatis, M. scrofulaceum, M. xenopi, M. kansasii, M. simiae and M. genavense. Rev Sci Tech 20(1): 265–290.
Bertelsen MF, Grondahl C, Giese SB (2006): Disseminated Mycobacterium celatum infection in a white-tailed trogon (Trogon viridis). Avian Pathol 35(4): 316–319.
Biet F, Boschiroli ML, Thorel MF, Guilloteau LA (2005): Zoonotic aspects of Mycobacterium bovis and Mycobacterium avium-intracellulare complex (MAC). Vet Res 36(3): 411–436.
BirdLife International (2016): Tauraco erythrolophus. The IUCN Red List of Threatened Species 2016: e.T22688346A93193784. https://dx.doi.org/10.2305/IUCN.UK.2016-3.RLTS.T22688346A93193784.en (11. September 2020).
Bottcher D, Paar M, Wittenbrink MM, Muller K, Bischofberger L, Schoon HA (2016): Mycobacterium genavense Infection as a Cause of Disseminated Granulomatous Inflammation in a Horse. J Equine Vet Sci 39: 76–79.
Böttger EC, Hirschel B, Coyle MB (1993): Mycobacterium genavense sp. nov. Int J Syst Bacteriol 43(4): 841–843.
Brannian RE (1993): Diseases in turacos, go-away birds and plantain-eaters. In: Fowler ME (ed.), Zoo and Wild Animal Medicine – Current Therapy. Saunders, Philadelphia, 237–240.
Chevrier D, Oprisan G, Maresca A, Matsiota-Bernard P, Guesdon JL (1999): Isolation of a specific DNA fragment and development of a PCR-based method for the detection of Mycobacterium genavense. FEMS Immunol Med Microbiol 23(3): 243–252.
Cochard T, Branger M, Supply P, Sreevatsan S, Biet F (2020): MAC-INMV-SSR: a web application dedicated to genotyping members of Mycobacterium avium complex (MAC) including Mycobacterium avium subsp. paratuberculosis strains. Infect Genet Evol 77: 104075.
Cornelis G, Reynders M, Deprez J, Vankeerberghen A, Orlent H (2018): Disseminated Mycobacterium genavense infection in an immunocompetent adult: a case report. Clin Microbiol Infect 24(12): 1355–1356.
De Lorenzi G, Kamphuisen K, Biscontini G, Pacciarini M, Zanoni M, Luppi A (2018): Mycobacterium genavense Infection in a Domestic Ferret (Mustela putorius furo). Top Companion Anim Med 33(4): 119–121.
Dvorska L, Matlova L, Ayele WY, Fischer OA, Amemori T, Weston RT, Alvarez J, Beran V, Moravkova M, Pavlik I (2007): Avian tuberculosis in naturally infected captive water birds of the Ardeideae and Threskiornithidae families studied by serotyping, IS901 RFLP typing, and virulence for poultry. Vet Microbiol 119(2-4): 366–374.
Ferrer L, Ramis A, Fernandez J, Majo N (1997): Granulomatous dermatitis caused by Mycobacterium genavense in two psittacine birds. Vet Dermatol 8(3): 213–219.
Fischer O, Matlova L, Bartl J, Dvorska L, Melicharek I, Pavlik I (2000): Findings of mycobacteria in insectivores and small rodents. Folia Microbiol (Praha) 45(2): 147–152.
Friedrich-Loeffler-Institut (2019): Amtliche Sammlung von Verfahren zur Probenahme und Untersuchung von Untersuchungsmaterial tierischen Ursprungs im Hinblick auf anzeigepflichtige Tierseuchen. https://www.fli.de/de/publikationen/amtliche-methodensammlung/.
Gomez G, Saggese MD, Weeks BR, Hoppes SM, Porter BF (2011): Granulomatous Encephalomyelitis and Intestinal Ganglionitis in a Spectacled Amazon Parrot (Amazona albifrons) Infected with Mycobacterium genavense. J Comp Pathol 144(2-3): 219–222.
Hoop RK, Bottger EC, Ossent P, Salfinger M (1993): Mycobacteriosis Due to Mycobacterium genavense in 6 Pet Birds. J Clin Microbiol 31(4): 990–993.
Hoop RK, Bottger EC, Pfyffer GE (1996): Etiological agents of mycobacterioses in pet birds between 1986 and 1995. J Clin Microbiol 34(4): 991–992.
Huynh M, Pingret JL, Nicolier A (2014): Disseminated Mycobacterium genavense Infection in a Chinchilla (Chinchilla lanigera). J Comp Pathol 151(1): 122–125.
Iwamoto T, Nakajima C, Nishiuchi Y, Kato T, Yoshida S, Nakanishi N, Tamaru A, Tamura Y, Suzuki Y, Nasu M (2012): Genetic diversity of Mycobacterium avium subsp. hominissuis strains isolated from humans, pigs, and human living environment. Infect Genet Evol 12(4): 846–852.
Keymer IF, Jones DM, Pugsley SL, Wadsworth PF (1982): A survey of tuberculosis in birds in the Regent’s Park gardens of the Zoological Society of London. Avian Pathol 11(4): 563–569.
Kik MJ, Houwers DJ, Dinkla A (2010): Mycobacterium intermedium granulomatous pneumonia in a green oropendola (Psarocolius viridis). Vet Rec 167(6): 219–220.
Krause KJ, Reavill D, Weldy SH, Bradway DS (2015): Mycobacterium genavense in an African Penguin (Spheniscus Demersus). J Zoo Wildlife Med 46(4): 971–975.
Lahiri A, Kneisel J, Kloster I, Kamal E, Lewin A (2014): Abundance of Mycobacterium avium ssp. hominissuis in soil and dust in Germany – implications for the infection route. Lett Appl Microbiol 59(1): 65–70.
Manarolla G, Liandris E, Pisoni G, Sassera D, Grilli G, Gallazzi D, Sironi G, Moroni P, Piccinini R, Rampin T (2009): Avian mycobacteriosis in companion birds: 20-year survey. Vet Microbiol 133(4): 323–327.
Martin G, Schimmel D (2000): Die Mycobacterium avium-Infektion des Geflügels – (k)eine Gefahr für die menschliche Gesundheit? Dtsch Tierarztl Wochenschr 107(2): 53–58.
Mendenhall MK, Ford SL, Emerson CL, Wells RA, Gines LG, Eriks IS (2000): Detection and differentiation of Mycobacterium avium and Mycobacterium genavense by polymerase chain reaction and restriction enzyme digestion analysis. J Vet Diagn Invest 12(1): 57–60.
Möbius P, Lentzsch P, Moser I, Naumann L, Martin G, Köhler H (2006): Comparative macrorestriction and RFLP analysis of Mycobacterium avium subsp. avium and Mycobacterium avium subsp. hominissuis isolates from man, pig, and cattle. Vet Microbiol 117(2-4): 284–291.
Montali RJ, Bush M, Thoen CO, Smith E (1976): Tuberculosis in captive exotic birds. J Am Vet Med Assoc 169(9): 920–927.
Morita Y, Maruyama S, Hashizaki F, Katsube Y (1999): Pathogenicity of Mycobacterium avium complex serovar 9 isolated from painted quail (Excalfactoria chinensis). J Vet Med Sci 61(12): 1309–1312.
O.I.E. (2018): Chapter 3.3.6 – Avian Tuberculosis. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2019, Paris, France.
Patino WLC, Monge O, Suzan G, Gutierrez-Espeleta G, Chaves A (2018): Molecular Detection of Mycobacterium avium avium and Mycobacterium genavense in Feces of Free-living Scarlet Macaws (Ara macao) in Costa Rica. J Wildl Dis 54(2): 357–361.
Peat L (2017): EAZA Turaco Best Practice Guidelines. Louise Peat, Cotswold Wildlife Park.
Peters M, Prodinger WM, Gummer H, Hotzel H, Mobius P, Moser I (2007): Mycobacterium tuberculosis infection in a blue-fronted amazon parrot (Amazona aestiva aestiva). Vet Microbiol 122(3-4): 381–383.
Pfeiffer W, Braun J, Burchell J, Witte CL, Rideout BA (2017): Whole-genome analysis of mycobacteria from birds at the San Diego Zoo. PLoS One 12(3): e0173464.
Portaels F, Realini L, Bauwens L, Hirschel B, Meyers WM, de Meurichy W (1996): Mycobacteriosis caused by Mycobacterium genavense in birds kept in a zoo: 11-year survey. J Clin Microbiol 34(2): 319–323.
Radomski N, Thibault VC, Karoui C, de Cruz K, Cochard T, Gutierrez C, Supply P, Biet F, Boschiroli ML (2010): Determination of Genotypic Diversity of Mycobacterium avium Subspecies from Human and Animal Origins by Mycobacterial Interspersed Repetitive-Unit-Variable-Number Tandem-Repeat and IS1311 Restriction Fragment Length Polymorphism Typing Methods. J Clin Microbiol 48(4): 1026–1034.
Sanchez FD, Yela IJ, Alfonseca E, Campuzano J, Morales E, Aguilar C (2016): Respiratory tract infection caused by Mycobacterium bovis in a black swan (Cygnus atratus). Avian Pathol 45(1): 126–131.
Scherrer S, Landolt P, Carroli N, Stephan R (2018): Molecular Characterization of Mycobacterium avium subsp. hominissuis of Two Groups of Lymph Nodes, Being Intradermal Tuberculin or Interferon-Gamma Test Positive and Negative, Isolated from Swiss Cattle at Slaughter. Front Vet Sci 5: 32.
Schrenzel M, Nicolas M, Witte C, Papendick R, Tucker T, Keener L, Sutherland-Smith M, Lamberski N, Orndorff D, Heckard D, Witman P, Mace M, Rimlinger D, Reed S, Rideout B (2008): Molecular epidemiology of Mycobacterium avium subsp. avium and Mycobacterium intracellulare in captive birds. Vet Microbiol 126(1-3): 122–131.
Shitaye EJ, Grymova V, Grym M, Halouzka R, Horvathova A, Moravkova M, Beran V, Svobodova J, Dvorska-Bartosova L, Pavlik I (2009): Mycobacterium avium subsp. hominissuis infection in a pet parrot. Emerg Infect Dis 15(4): 617–619.
Shitaye JE, Halouzka R, Svobodova J, Grymova V, Grym M, Skoric M, Fictum P, Beran V, Slany M, Pavlik I (2010): First isolation of Mycobacterium genavense in a blue headed parrot (Pionus menstruus) imported from Surinam (South America) to the Czech Republic: a case report`. Vet Med (Praha) 55(7): 339–347.
Smit T, Eger A, Haagsma J, Bakhuizen T (1987): Avian tuberculosis in wild birds in the Netherlands. J Wildl Dis 23(3): 485–487.
Stamper A, Norton T, Loomis M (1998): Acid-fast bacterial infection in four turacos. J Avian Med Surg 12(2): 108–111.
Starkova DA, Otten TF, Mokrousov IV, Viazovaia AA, Vishnevskii BI, Narvskaia OV (2013): [Genotypic characteristics of Mycobacterium avium subsp. hominissuis strains]. Genetika 49(9): 1048–1054.
Steinmetz HW, Rutz C, Hoop RK, Grest P, Bley CR, Hatt JM (2006): Possible human-avian transmission of Mycobacterium tuberculosis in a green-winged macaw (Ara chloroptera). Avian Dis 50(4): 641–645.
Tell LA, Woods L, Cromie RL (2001): Mycobacteriosis in birds. Rev Sci Tech 20(1): 180–203.
Tell LA, Leutenegger CM, Larsen RS, Agnew DW, Keener L, Needham ML, Rideout BA (2003): Real-time polymerase chain reaction testing for the detection of Mycobacterium genavense and Mycobacterium avium complex species in avian samples. Avian Dis 47(4): 1406–1415.
Theuss T, Aupperle H, Eulenberger K, Schoon HA, Richter E (2010): Disseminated Infection with Mycobacterium genavense in a Grizzled Giant Squirrel (Ratufa macroura) Associated with the Isolation of an Unknown Mycobacterium. J Comp Pathol 143(2-3): 195–198.
Thibault VC, Grayon M, Boschiroli ML, Hubbans C, Overduin P, Stevenson K, Gutierrez MC, Supply P, Biet F (2007): New variable-number tandem-repeat markers for typing Mycobacterium avium subsp. paratuberculosis and M. avium strains: comparison with IS900 and IS1245 restriction fragment length polymorphism typing. J Clin Microbiol 45(8): 2404–2410.
Tocidlowski ME (2012): Musophagiformes. In: Miller ER, Fowler ME (eds.), Fowler’s Zoo and Wild Animal Medicine. Elsevier Saunders, St. Louis, Missouri, 206–209.
Tran QT, Han XY (2014): Subspecies identification and significance of 257 clinical strains of Mycobacterium avium. J Clin Microbiol 52(4): 1201–1206.
Turenne CY, Wallace R, Jr., Behr MA (2007): Mycobacterium avium in the postgenomic era. Clin Microbiol Rev 20(2): 205–229.
Ullmann LS, Dias-Neto RD, Cagnini DQ, Yamatogi RS, Oliveira JP, Nemer V, Teixeira RHF, Biondo AW, Araujo JP (2016): Mycobacterium genavense infection in two species of captive snakes. J Venom Anim Toxins Incl Trop Dis 22: 27.
van Ingen J, Turenne CY, Tortoli E, Wallace RJ, Brown-Elliott BA (2018): A definition of the Mycobacterium avium complex for taxonomical and clinical purposes, a review. Int J Syst Evol Microbiol 68(11): 3666–3677.
Wayne LG, Good RC, Tsang A, Butler R, Dawson D, Groothuis D, Gross W, Hawkins J, Kilburn J, Kubin M, Schroder KH, Silcox VA, Smith C, Thorel MF, Woodley C, Yakrus MA (1993): Serovar Determination and Molecular Taxonomic Correlation in Mycobacterium avium, Mycobacterium intracellulare, and Mycobacterium scrofulaceum – a Cooperative Study of the International Working Group on Mycobacterial Taxonomy. Int J Syst Bacteriol 43(3): 482–489.
Wilson SC, Carpenter JW (1994): Investigation of Suspected Mycobacteriosis in a Group of Tropical Birds at the Topeka Zoological Park. Proc Am Assoc Zoo Vet, 164–167.
Witte CL, Hungerford LL, Papendick R, Stalis IH, Rideout BA (2008): Investigation of characteristics and factors associated with avian mycobacteriosis in zoo birds. J Vet Diagn Invest 20(2): 186–196.


Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2020-39-Barth.pdf (0.24 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2020-39-Barth-Tabelle1.pdf (0.2 MB) herunterladen möchten



{kind=link}
{kind=link}
{kind=link}