
ASP-Bekämpfung: Kadaversuche aus der Luft – Möglichkeiten und Einschränkungen
Berliner und Münchener Tierärztliche Wochenschrift 134, 1-14
DOI: 10.2376/1439-0299-2020-46
© Schlütersche Fachmedien GmbH. 2021
Eingereicht: 11. Januar 2021
Akzeptiert: 29. Juni 2021
Publiziert: 08/2021
Summary
The removal of wild boar carcasses is one of the first important countermeasures after an ASF outbreak. Thermal imagers that visualise body temperature are already in use for selective carcass detection. In addition to carrion-specific infrared radiation (IR-radiation) derived from natural body temperature or decay heating, non-carrion specific heat sources such as the absorption of sunlight, limit detection selectivity. To understand the influence of different heat sources on the IR-radiation phenology of carcasses better, we monitored the internal temperature of nine culled wild boar (Sus scrofa) during decomposition using dummies as a control. Culling took place between June and November and the objects were placed in fenced-off areas. Some of them were exposed to direct sunlight throughout, while others remained in the shade. For detection, we used both stationary and helicopter mounted thermal imagers. As expected, all culled boar had cooled down approximately two days after death. In summer, in culled boar that were placed at ambient temperatures above 10 °C, decay-heating started after 3–7 days and lasted up to four weeks. In the carcasses that were exposed to direct sunlight, absorption had a similar effect on IR-radiation as is normally observed from internal heating. In boar culled during November–December at an ambient temperature below 10 °C, we found no signs of decomposition or decay-heating. Solar absorption was the main source of IR-radiation after algor mortis. We therefore conclude that thermal imaging will be less efficient for detecting carcasses during colder seasons (Ø <10 °C; November–March) and at other times of the year on sunny days and the subsequent nights following such days. Considering the highlighted challenges relating to the detection of wild boar carcasses using IR-radiation, and the fact that moribund boar prefer places with IR-shielding vegetation to die, the continued reliance on a ground search approach for detecting infected carrion is still inevitable at present.
Zusammenfassung
Im ASP-Management ist die Kadaverbeseitigung eine der ersten Gegenmaßnahmen. Dabei wird der Einsatz von Infrarotkameras zur Detektion der Körperwärme bereits für eine selektive Kadaversuche praktiziert. Doch neben kadaverspezifischer Wärmestrahlung aufgrund Restkörper- oder Verwesungswärme können kadaverunspezifische Wärmequellen wie die Absorption von Sonnenlicht die Selektivität einschränken. Um den Einfluss der verschiedenen Wärmequellen auf die IR-Abstrahlungsphänologie eines Kadavers besser zu verstehen, wurden neun zwischen Juni und November getötete Wildschweine (Sus scrofa) sowie Wildschweinattrappen in umzäuntem Gelände ausgelegt. Ein Teil wurde der direkten Sonneneinstrahlung ausgesetzt. Zur Detektion wurden IR-Kameras an einem Hubschrauber sowie stationäre Anlagen genutzt. Wie zu erwarten, kühlten alle Versuchstiere am zweiten Tag nach der Tötung vollständig aus. Die Sommerkadaver bei Umgebungstemperaturen von > 10 °C zeigten nach dem dritten bis siebten Tag Verwesungswärme, die bis zu vier Wochen anhielt. Die Herbstkadaver bei Umgebungstemperaturen von < 10 °C zeigten weder Zeichen von Verwesung noch von Verwesungswärme. Im Freien heizte die Absorption von Sonnenlicht die Kadaver oder Attrappen ähnlich auf wie der Verwesungsmetabolismus. Bei den Herbstkadavern war Absorption die einzige nennenswerte Wärmequelle nach Algor mortis. Die Effizienz von Infrarotkameras zur selektiven Detektion von Kadavern wird daher in der kalten Jahreszeit (Oktober/November bis März/April) und ganzjährig während sonniger Wetterphasen und in der Folgenacht eingeschränkt sein. Da sich kranke Wildschweine bevorzugt in Bereiche mit IR-abschirmender Vegetation zurückziehen, sind bodengestützte Kadaversuchstrategien im ASP-Management unumgänglich.
Introduction
Within an African Swine Fever (ASF) outbreak area, most wild boar get infected by direct contact with dead conspecifics that carry the virus (Chenais et al. 2019). Carcasses can facilitate the spread of the disease for several weeks to months during decomposition (Miteva et al. 2020, Zani et al. 2020). Thus, the rapid removal of wild boar carcasses during a sylvatic ASF outbreak is one of the most effective and important countermeasures (Miteva et al. 2020). However, the rapid detection of the carcasses remains a challenging task as wild boar (Sus scrofa) prefer dense vegetation during resting time and moribund boar infected with ASF frequently die in areas that are poorly accessible (Morelle et al. 2019). Usually, search teams on the ground try to locate boar carcasses, either visually or with the aid of trained detection dogs. Both approaches are time-consuming and expensive. On the other hand, it is known from forensic research that both human bodies and domestic pigs (Sus scrofa domesticus) that are used as human body models for decomposition studies can produce a considerable amount of heat during decomposition (Johnson et al. 2013, Lee et al. 2018, Sharanowski et al. 2008). The generation of heat during decomposition is the result of necrophagous insect larvae and bacterial activity inside the carcass (DesMarais 2014). Depending on ambient conditions, the decomposition process can last up to three to four weeks, and results in a net difference of body surface temperature to ambient temperature of up to 10 °C or more. Surface temperature can be measured non-invasively using infrared or thermal imagers (DesMarais 2014, Edelman et al. 2013, Johnson et al. 2013). Thermal imaging cameras mounted on an aircraft (e.g. a drone or helicopter) can make use of this temperature difference to locate decomposing carcasses from a distance (Amendt et al. 2017, Bodnar et al. 2019, Butters et al. 2020, Descalzi 2019, Lee et al. 2018, Murray et al. 2018). Research has shown that decaying clothed bodies and mammalian carcasses wrapped in plastic or carpet, or even buried, still produce internal heat to generate sufficient surface temperature (difference >0,1 °C) contrasts to the background (Butters et al. 2020, Kelly 2006, Lee et al. 2018, McDaneld 2016). We therefore assume that a decaying wild boar carcass, despite being covered with thick, highly insulating fur, would be detectable using infrared or thermal imagers during putrefaction. However, many other natural objects have similar abilities to emit infrared radiation (IR-radiation) with emissivity values and absorption coefficients for electromagnetic radiation (ability to weaken the intensity of electromagnetic radiation passing an object) comparable with those of skin or fur (Edelman et al. 2013).
These objects such as bare soil, wooden material, dried-out vegetation or stones could emit thermal radiation within the range of a carcass. We wondered to which extend the absorption of sunlight contributes to the net IR-radiation of a carcass beside any internal heat sources.
Moreover, in case of a given similar heat capacity and thermal conductivity, many such objects emit IR-radiation even hours after sunset, mimicking the properties of a warm, decaying carcass into night time. In colder periods of slowed down – or even blocked – activity of necrophagous insects and bacteria (Anderson 2000, Kelly 2006, Lee et al. 2018), these abiotic mechanisms could determine the thermal signature of a carcass only.
Top Job:
As the efficiency of carcass detection requires a reliable differentiation of objects based on the emitted IR-radiation, these masking effects should be considered. To gain a better understanding of such interfering effects, this study aims to differentiate physical (external) warming from decomposing (internal) warming by means of an experimental set up using wild boar carcasses and dummies exposed to direct sunlight or shade during summer and winter.
Materials and methods
We used nine culled wild boar with an average life weight of 28 kg (range of 20–40 kg, details in Table 3). We also prepared wild boar dummies to have the thermal conductivity and absorption properties similar to a wild boar carcass but excluding the decay metabolism as an energy source. A wild boar dummy is made up of a tanned wild boar skin wrapped around water tubes each filled with approximately 25 l of tap water.
The experimental (culled wild boar) and control (wild boar dummy) objects were placed in different fenced-off areas and arranged in five different experimental settings at different times of the year. Three of the experimental settings were in summer (June–August/September) and two were in winter (November/December) as shown below:
- Setting 1, Winningen (Rhineland-Palatinate, Germany): During the first trial in summer (June 2019), two wild boar carcasses were positioned side by side in the open (Fig. 1).
- Setting 2, Winningen (Rhineland-Palatinate, Germany): In the second trial in summer (August 2019), a wild boar dummy was placed in the open as a control next to a recently opened and exposed wild boar carcass.
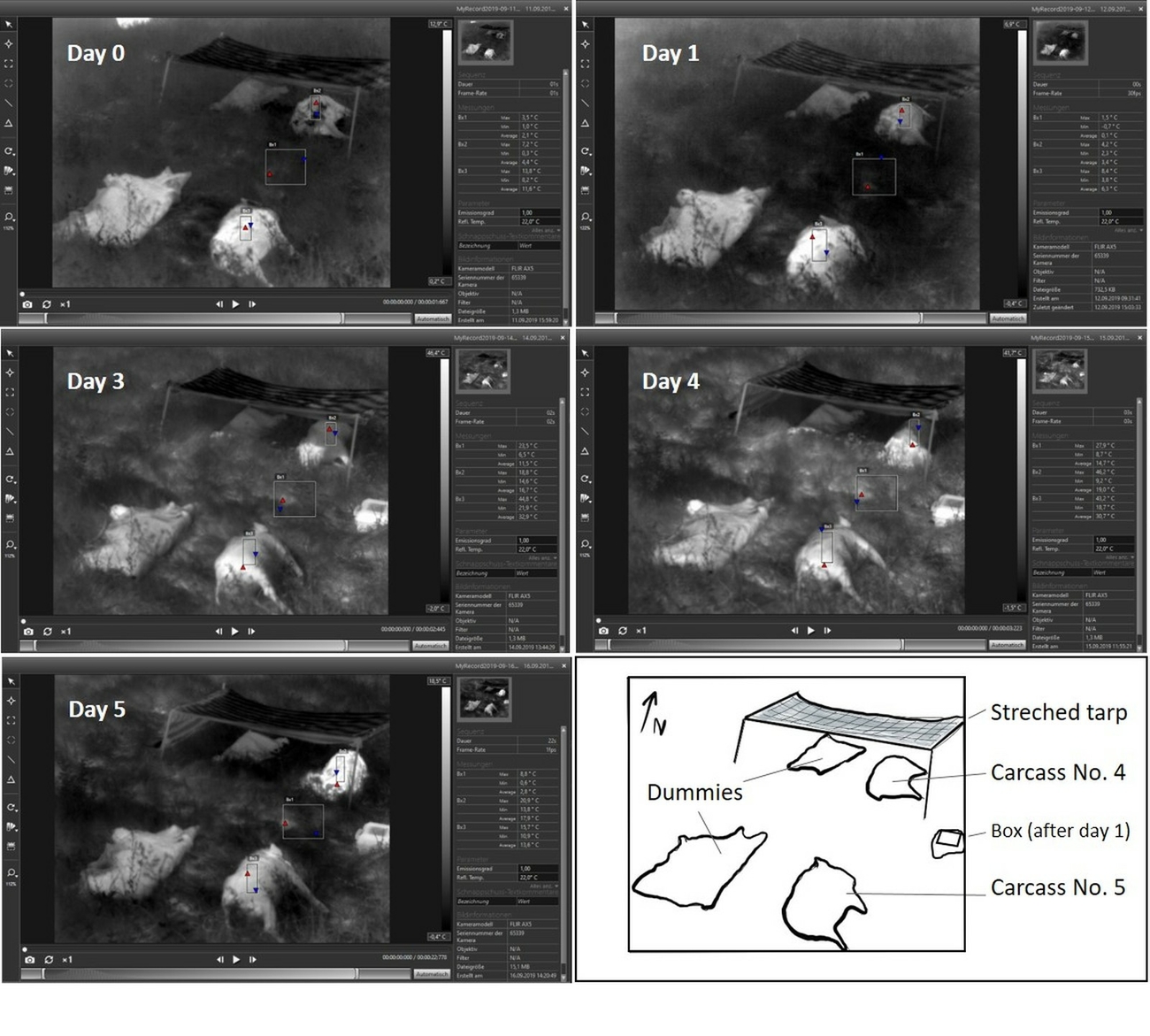
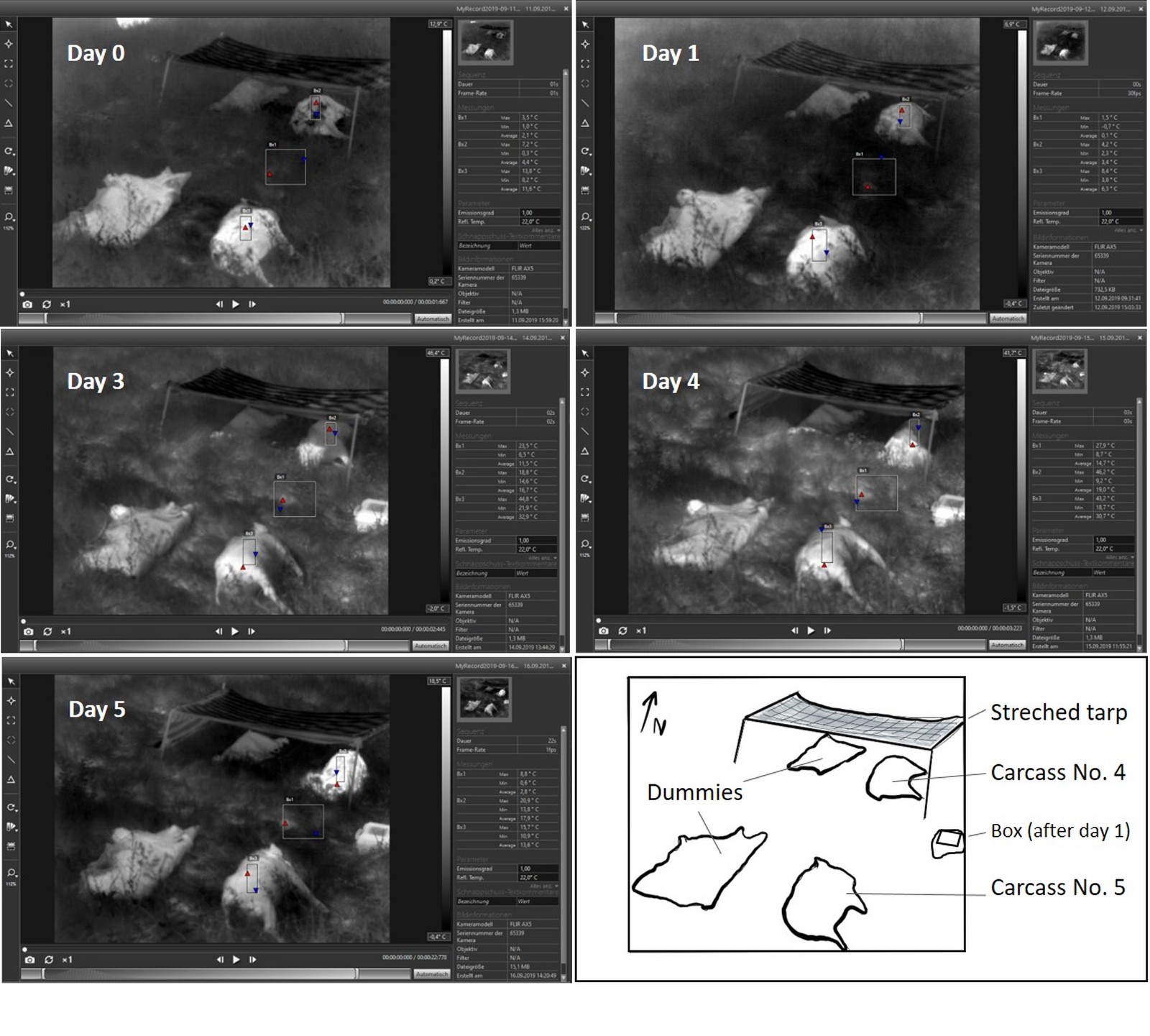
- Setting 3, Bassenheim (Rhineland-Palatinate, Germany): In late summer (September 2019), a wild boar carcass/dummy pair was placed in the open and another pair was positioned nearby in constant shade (generated with a self-made sun shading made up of stretched tarp; Fig. 2).
- Setting 4, Hochscheid (Rhineland-Palatinate, Germany): In early winter (November 2019), a wild boar carcass/dummy pair was again positioned in the open and another pair nearby in constant shade.
- Setting 5, Hochscheid (Rhineland-Palatinate, Germany): In winter (December 2019) in a similar set-up as described in Setting 4 with the only difference being large lacerations due to penetrating gunshot wounds.
All wild boar, except for one animal in Setting 1, were kept in a semi-natural enclosure before culling. To avoid any impacts on insect activity and natural decomposition processes, we did not use drugs to euthanize study animals. Most of the animals were culled by means of a close-range head shot to limit the number of artificial body openings resulting from penetrating gunshot wounds. Only one animal in Setting 1 and the animals in Setting 5 were culled with a typical hunting-distance shot to the chest that produced a large penetrating gunshot wound.
All wild boar carcasses were exposed to ambient conditions in fenced-off areas to prevent interference by foxes or other ground scavengers. Airborne scavengers such as buzzards (Buteo buteo) had unlimited access to the carcasses but were rarely observed in the study areas.
Measurement of wild boar carcass and wild boar dummy internal temperatures
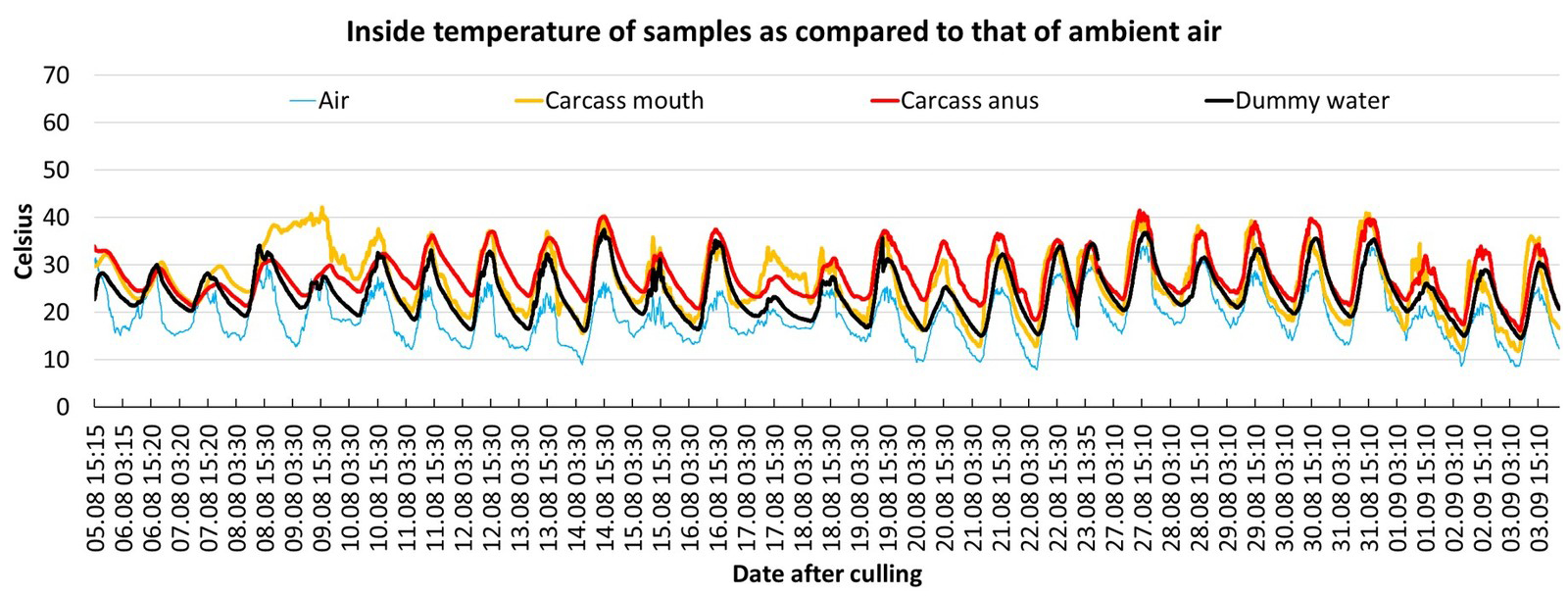
We measured and stored internal sample temperature with a temperature-measuring probe (Trueber T100) and a data logger. Temperature measurements were taken at least every hour, starting after culling and lasting for three to four weeks (Fig. 4). Temperature probes were placed 20 cm deep in the mouth and anus or under the skin of the wild boar carcasses. In case of the dummies, the temperature probes were located between the skin and the water tubes. The ambient air temperature was measured in a box placed 50 cm above ground in the shade (Fig. 1). To simplify the data capture, we measured the daily minimum and maximum ambient temperatures of each carcass and dummy, and calculated the daily average from these results.
IR-radiation
Thermal images were taken with a FLIR camera system to visualize the differences in IR-radiation between the carcasses, the dummies, and their surroundings. The images captured are in greyscale, with colder areas being shown as darker. A FLIR-System (Star SAFIRE® 380-HDC) was attached to a helicopter (Polizei-Hubschrauberstaffel Winningen, Rhineland-Palatinate), which flew over the study area 32 times (Settings 1 and 2 were observed from 16 June to 3 September 2019 with 56% of the images being captured at night). A ground station (FLIR A65) was positioned at a height of 2–4 m above ground and a distance of 8–9 m to the objects (Settings 3, 4 and 5) from 11 September to 15 December 2019. The FLIR A65 is a thermal imaging temperature sensor that provides linear output of recorded temperatures. When the emissivity values are similar, this technique can be used as a proxy of temperature differences between observed objects. Surface temperature differences were taken between the ground and objects in Setting 3. Due to handling errors during the evaluations in Setting 3, we could only store six measurements taken between 11–16 September 2019. In Settings 4 and 5, surface temperature differences were taken between the wild boar dummy and the carcasses in the shade because ground measurements failed.
Daily weather data (number of hours of sunshine and air temperature) were collected from data provided by the German National Meteorological Service (Deutscher Wetterdienst; https://www.dwd.de/) station ‘Andernach’, which is situated 13 km from Settings 1 and 2, and 6 km from Setting 3, or from the meteorological station ‘Hahn’, which is situated 6 km from Settings 4 and 5.
Results
Carcass temperatures recorded in summer (June–September 2019; Settings 1 and 3)
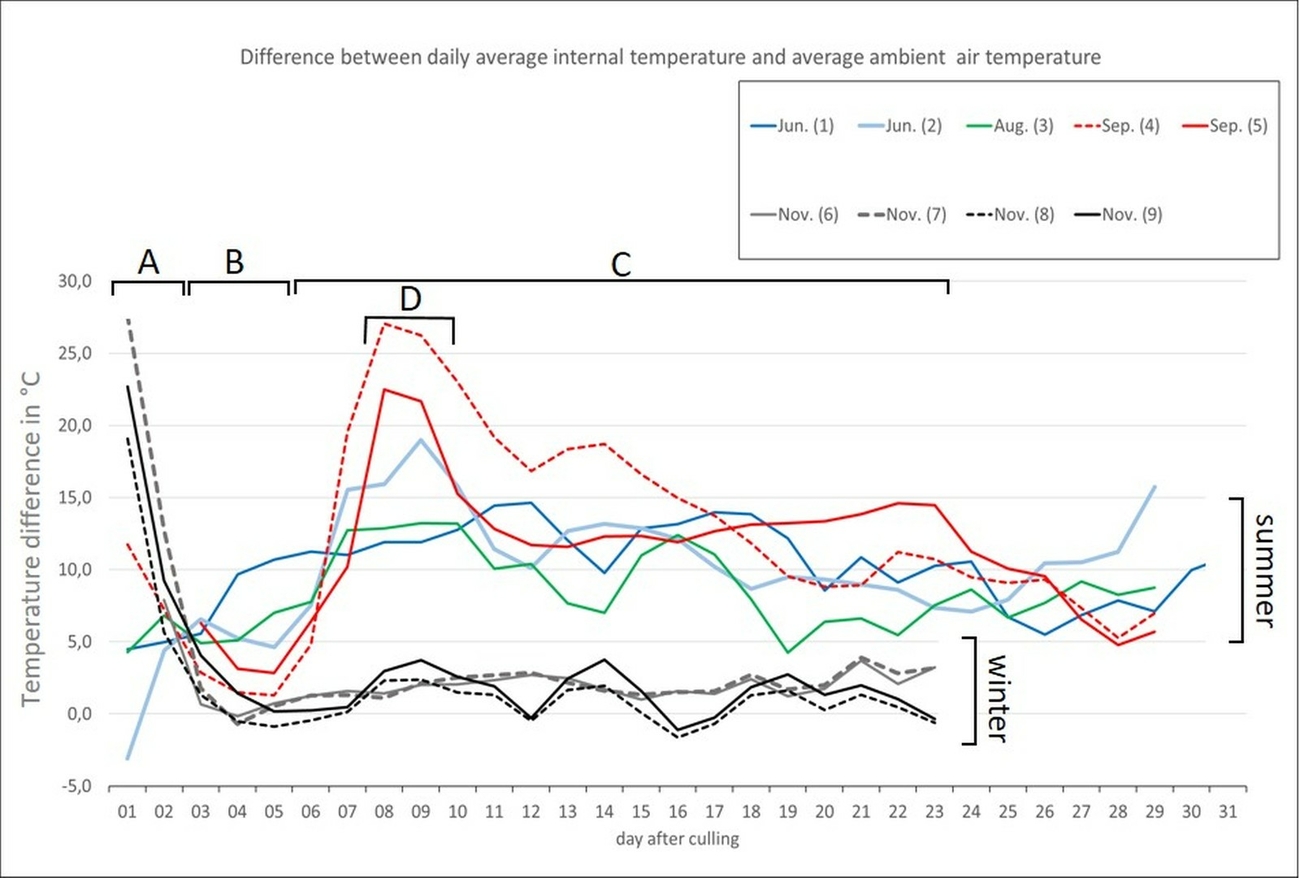
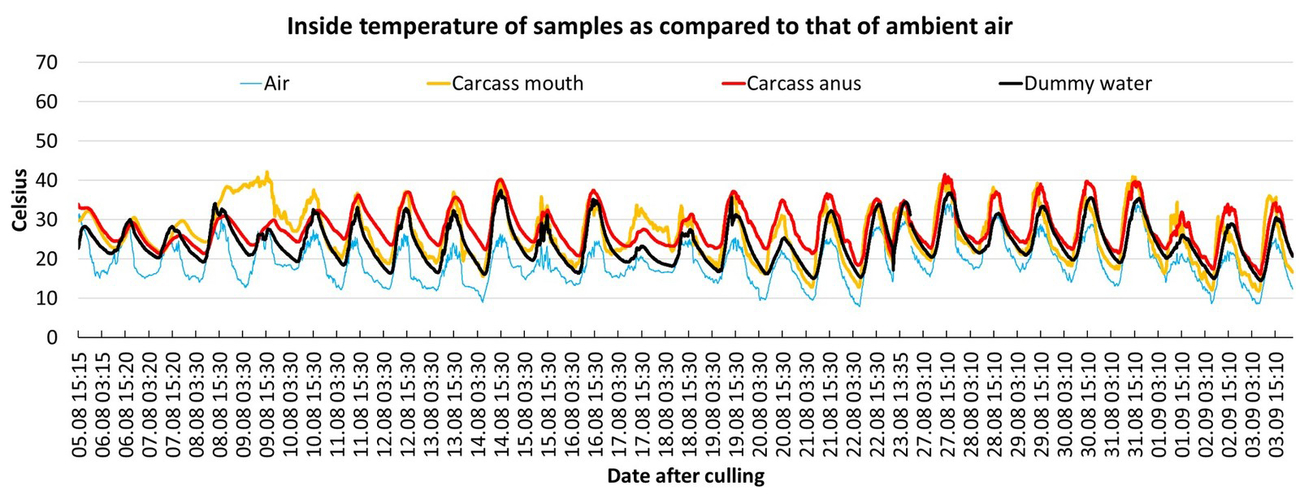
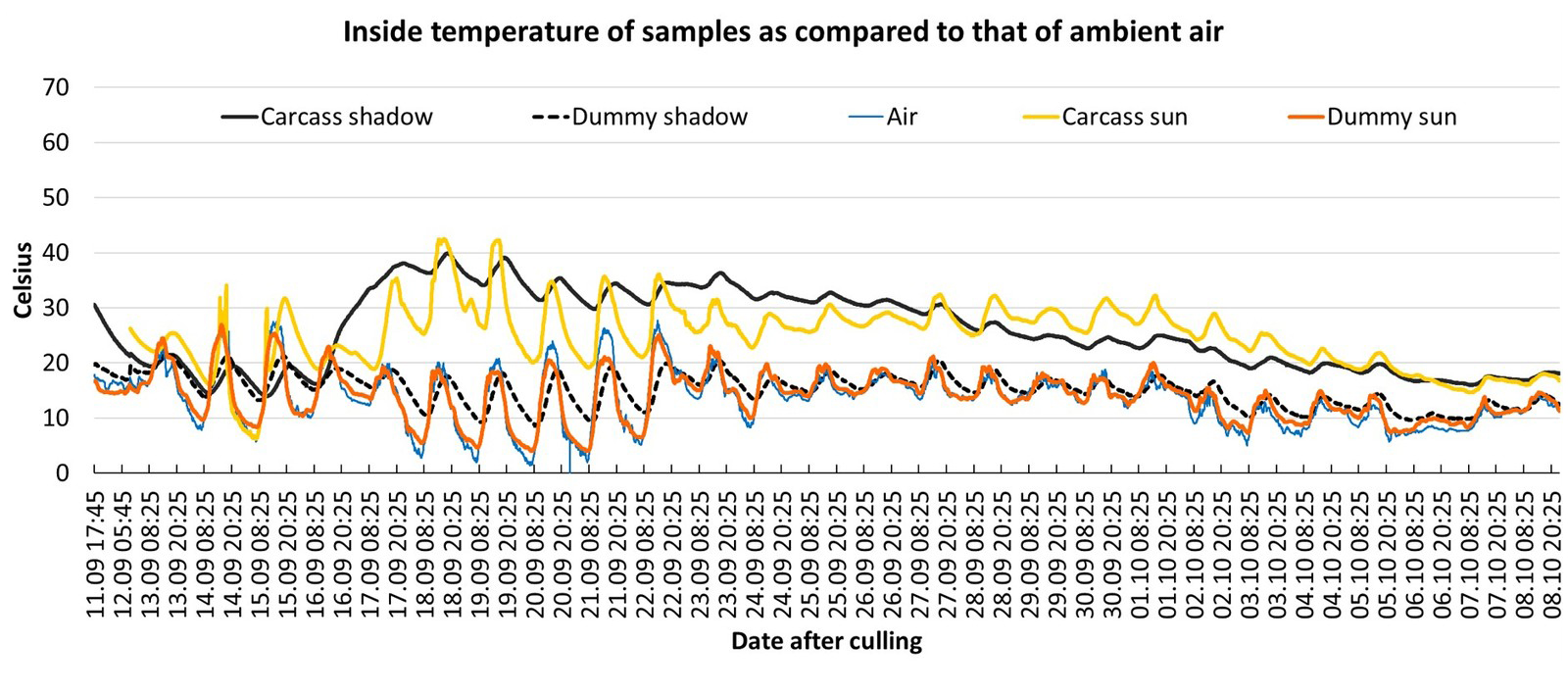
During the first two days after culling, all carcasses cooled down at a rate of approximately 0,5–1 °C per hour (ambient temperatures ranged from 0–17 °C). Due to ambient conditions, the differences between the ambient air temperature and that of the carcasses could be positive or negative (Period A in Fig. 4). From day 3 to day 7, all four summer carcasses (mean daily average air temperature of 14–21 °C) became warmer than the ambient air temperature for a period of between 12–21 days after culling (Table 1). During this period, the daily average temperature of the carcasses was mostly found to be between 9–18 °C above the daily average ambient air temperature (Table 1). For some days, the temperature difference between the ambient air and the carcass could be even higher (Period B/C in Fig. 4). Especially during periods of colder weather (Fig. 6, period between 18–21 September 2019), the carcasses showed the highest temperature difference to the ambient air temperature of up to 26 °C (Period D in Fig. 4). Period C represents the timeframe during which the most pronounced maggot mass was visible in and around the carrion (Fig. 3). All measured temperatures in the carcasses showed a diurnal increase and decrease, following a day-night rhythm (Fig. 5 and Fig. 6). The internal temperature of the summer carcass in the shade (Setting 3, animal 4) displayed a similar pattern, but with less fluctuation than the carcasses that were exposed in the open (Setting 3, animal 5; Table 1, Fig. 6). Both the internal and skin temperature of the carcasses correlated more directly with the duration of sunshine per day (R2 = 0.3) than with the maximum air temperature (R2 = 0.02).
Carcass temperatures recorded during winter (November–December 2019; Settings 4 and 5)
In contrast to the carcasses from the summer period, the winter carcasses (mean daily average air temperature of 4–5 °C) did not show any signs of decomposition, internal heating or any obvious infestation with the larvae of necrophagous insects for at least one month after culling (Fig. 4 and Fig. 7).
Wild boar dummy temperatures
In the case of the controls, summer and winter temperature measurements were also regarded separately.
In summer, considering the differences in the daily average temperature, the wild boar dummies were warmer than the ambient air, but cooler than active decomposing wild boar carcasses (Table 1). Although the dummies were coldest compared to the carcass when both were in the shade, the situation changed when they were exposed to direct sunlight. In August and September (in June/July no dummies were used), i.e. during periods characterised by pronounced internal decay heating and sunshine, the temperatures measured in the dummy could be within the temperature range of the carcass. In summer, this was the case between 21–24 August 2019 with more than 12 hours of sunshine being recorded (Fig. 5). During this period, the dummy internal temperature was almost equal to that of the decaying carcass. In autumn, between 21–22 September 2019, with 11 and 6 hours of sunshine the dummy maximum internal temperatures reached the levels of the carcass minimal internal temperatures (Fig. 6).
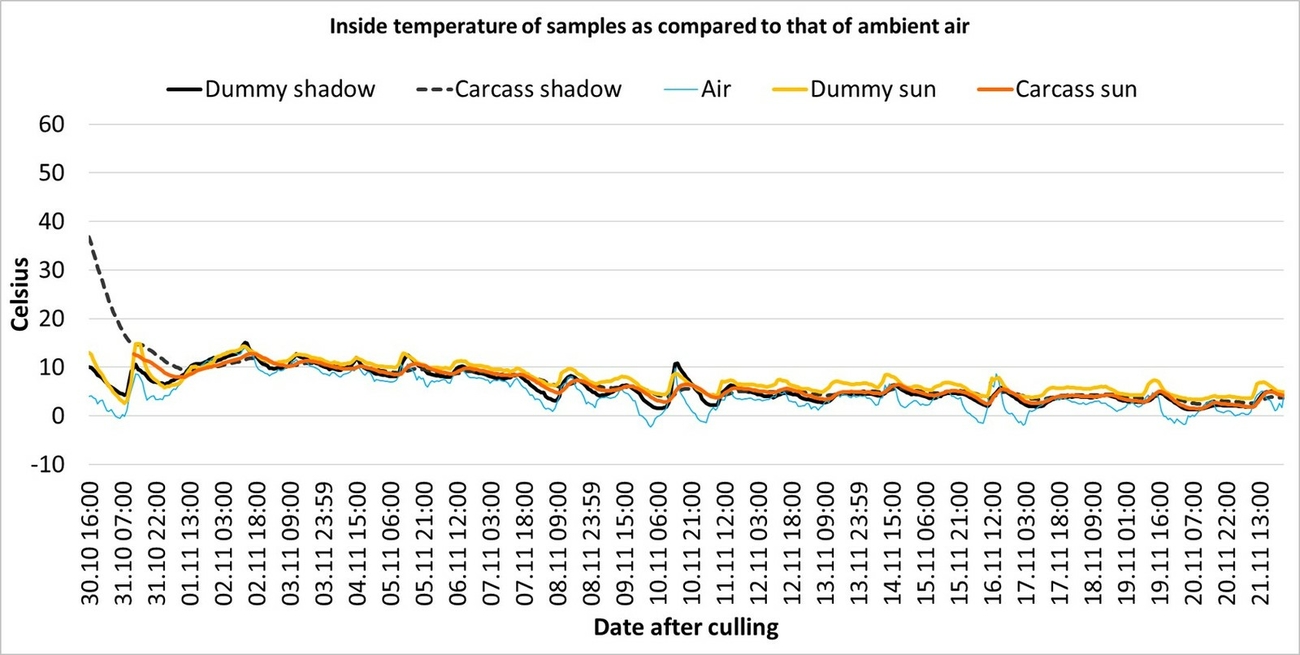
In winter (Settings 4 and 5), the temperature profiles of the dummies and carcasses were similar and close to that of the ambient air temperature, regardless of whether they were exposed to sunlight or not (Fig. 4 and 7).
IR-radiation
In general, thermal images captured by using the FLIR-cameras could visualize the carcasses when compared to the surrounding objects. This was the case up until two days after culling, as long as algor mortis was not completed, or after internal decay heating started from day three to seven after culling. Interestingly, even carcasses older than 50 days were still detectable. However, if carcasses were exposed in the open, especially during periods with direct sunlight, differentiation of objects became difficult as the thermal imager showed similar IR-signatures (here similar greyscales) for surrounding objects and dummies (Fig. 8, 9, 10, 11). This phenomenon could last until the following night after a sunny day (Fig. 12).
The open exposed and shaded carcass/dummy pairs in Settings 3 gave more insight into these masking effects via absorption of sunlight (Fig. 13).
Summer period (August–September, Setting 3)
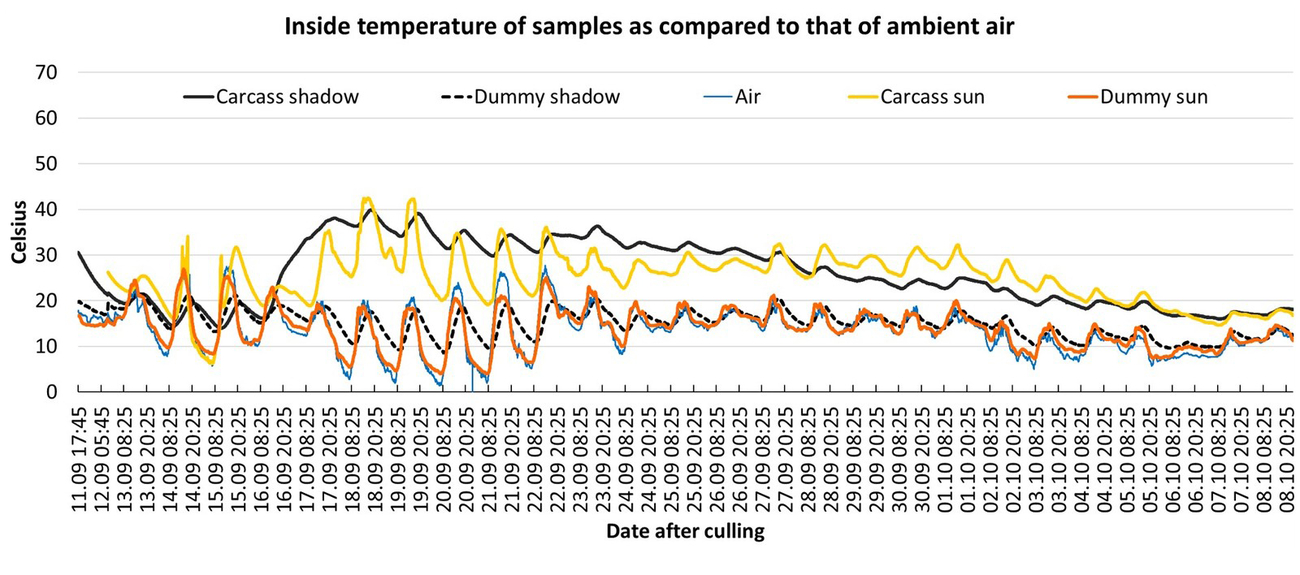
On day zero and one day after culling, both carcasses were warmer than the grassy ground. However, the carcass exposed to sunlight (approximately 4–5 hours sunshine; Table 2) was between 3–21 °C warmer than the carcass in the shade. The dummy in the open showed a similar IR-signature and only the dummy in the shade was colder.
On the third and fourth day after culling, algor mortis was completed, but decay heating had not yet started. This was a period characterised by consistently sunny and warm weather. The carcass and dummy exposed in the open reached temperatures of 9.5 °C higher than that of the ground temperature. In contrast, the carcass parts that were in the shade were 5.5 °C cooler than the exposed ground, and 20 °C cooler than the carcass exposed in the open. The dummy in the shade showed a similar ‘cold’ surface IR-signature whereas the dummy in the open showed a similar ‘warm’ IR-signature.
On day five, when internal heating started, the surface temperature of the carcass in the shade was 5 °C higher than that of the carcass exposed in the open, when compared to the ground surface temperature. This day was characterised by overcast skies. The dummy in the shade was warmer than the surface ground temperature (slightly lighter shade of grey), but this difference was less pronounced when compared to the carcass in the shade and to the exposed carcass and dummy.
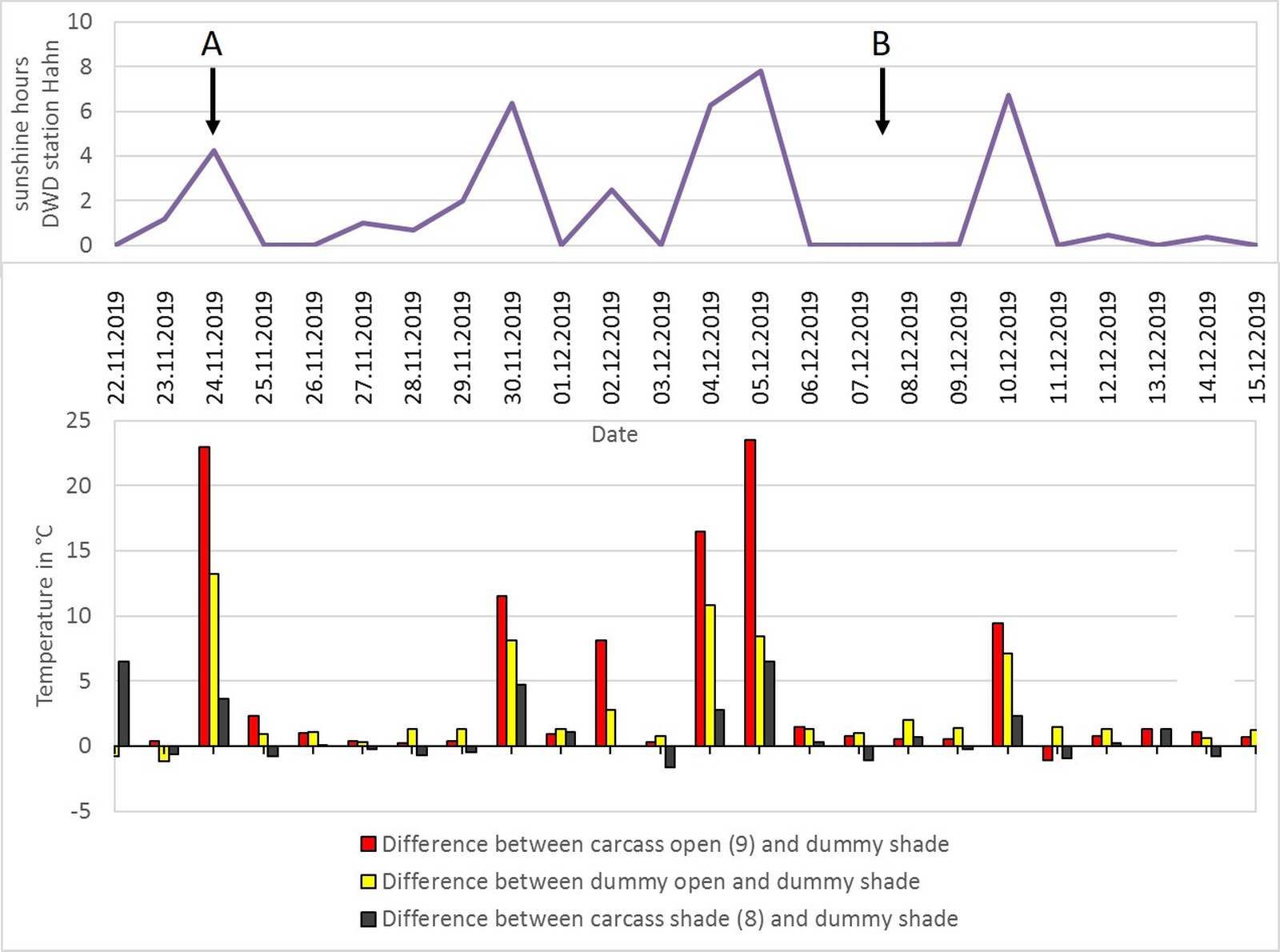
Winter period (November–December; Setting 4 and 5; Fig. 14)
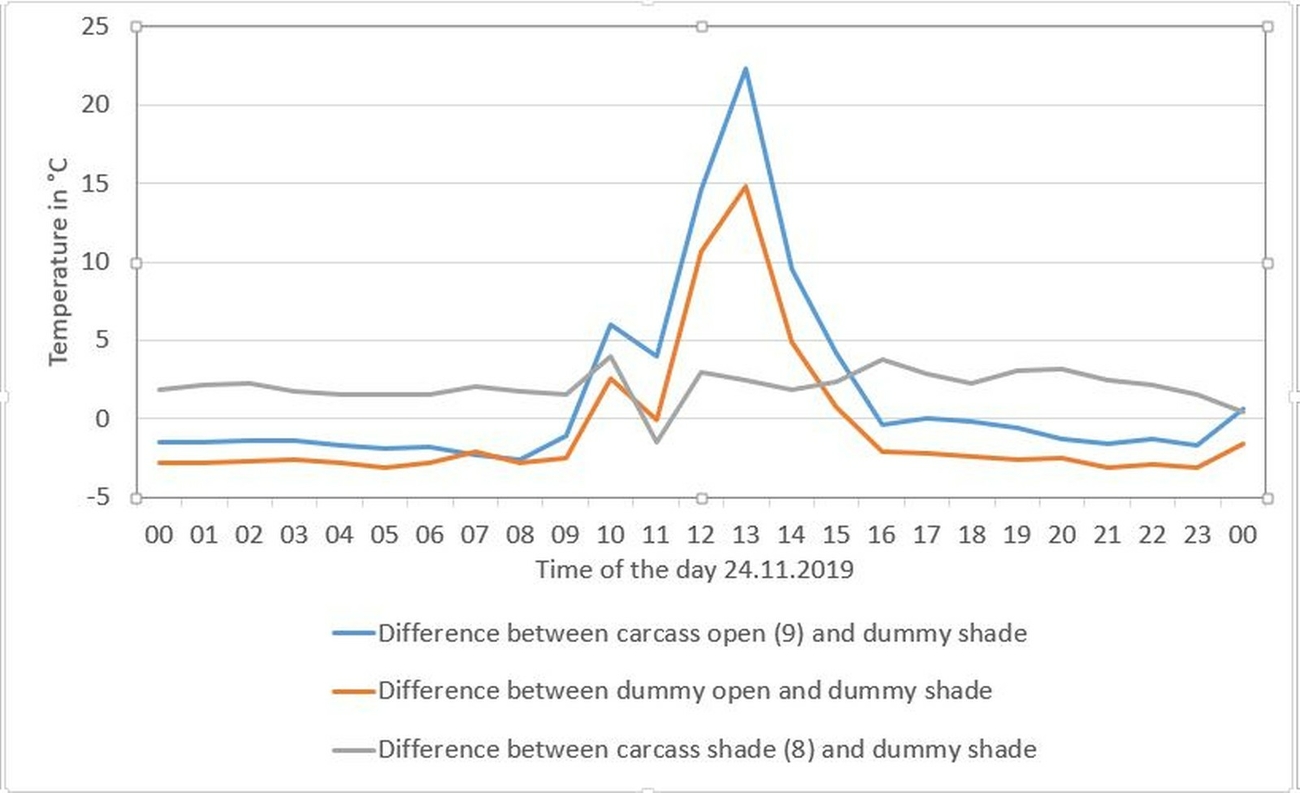
Only during the first two days both carcasses could be visualized in the thermal image (when compared to internal temperature in Fig. 4). Later, the carcass and dummy exposed in the open only reached temperatures that were 10–20 °C warmer than the shaded counterparts (e.g. Fig. 15; arrow A) during periods of sunshine. During periods that were overcast, only minor surface temperature differences could be observed between the objects exposed in the open and those in the shade (Fig. 15; arrow B). By taking a closer look at the hourly differences of the surface temperature between different objects on 24 November 2019 with a sunny period between 11 a.m. and 2 p.m., it is noticeable, that during the night hours, both the carcass and the dummy exposed in the open were colder than the shaded objects due to a higher negative radiation ratio (Fig. 16).
Discussion
As expected, the speed and phenology of decomposition in the medium-sized wild boar carcasses did not differ from that of similarly sized domestic pig carcasses (Amendt et al. 2017, DesMarais 2014, Johnson et al. 2013, Sharanowski et al. 2008). This study confirms that thermal imaging of wild boar carcasses could be a useful tool to detect carrion under natural conditions. Despite the fact that a wild boar carcass is covered with an insulating layer of fur, IR-emission was not hampered to such an extent as to limit detection by an infrared camera. However, the efficiency of such an approach is often limited in ASF management as thousands of hectares of suitable wild boar habitat must be covered (Tierseuchenbekämpfungsdienst Brandenburg). One concerning aspect is that masking effects of absorbed solar radiation generally decrease the detectability of carcasses in open areas. Indeed, our study demonstrated that absorbed solar radiation could dominate the IR-signature of a medium-sized mammalian carcass that is exposed in the open. With non-decaying dummies that have identical physical properties to wild boar carcasses in terms of their mass, emissivity, heat capacity, and thermal conductivity, we showed that these objects can be reheated in a similar way. In winter, when the decomposition process was inhibited, the absorption of sunlight was the only relevant energy source of IR-radiation for ground objects, including dead animals. Furthermore, due to heat capacity, an elevated IR-radiation of such items could last until the following night after a sunny day. Hence, abundantly occurring ground objects in a typical wild boar habitat such as bare ground, stones or wooden material can show IR-signatures that are similar or even identical to that of an exposed carcass, both during the day and at night. These masking effects might at least partly explain the limited efficiency of carcass detection strategies using the FLIR-technique, as a reliable IR-differentiation of objects is hampered by such effects.
Some recent studies, mostly with a forensic background, have examined the potential of drone- or helicopter-assisted infrared imaging techniques to facilitate the detection of corpses in large areas and at longer post-mortem intervals. In one of the first studies of this kind, Amendt et al. (2017) showed that internal decay heating is sufficient to detect pig carcasses from a helicopter several weeks after death. Murray et al. (2018) used drones to search for human remains and found that the surface temperature of desiccated skin can be changed by absorbing solar radiation. Lee et al. (2018) also worked with aircraft-mounted thermal imagers to measure the IR-radiation of the decomposing remains of eight decaying pigs at ambient temperatures ranging between 14–24 °C. The carcasses in this study remained several degrees warmer than the surroundings (on average), but detectability was shown to decline when the temperature contrast decreased under specific circumstances. Unfortunately, the authors did not mention the effect of solar radiation on thermal imaging. Descalzi (2019) also used thermal imagers mounted on drones for detecting larval aggregations associated with decomposing carcasses. She mentioned that masking effects were caused by decreased detectability due to sunlight, but without providing further details. In an Australian study, Butters et al. (2020) also found that IR-imagers operated from an unmanned aerial platform and used to search for decomposing remains were influenced by sunlight. Although some of these studies prolonged their measurements into the winter season, the local climate did not allow ambient temperatures to drop regularly below 10 °C. The development of necrophagous insect larvae was therefore merely slowed down, but a maggot mass still ensued (Butters et al. 2020, Kelly 2006, Lee et al. 2018). In this study, we demonstrated that no decay heating and consequently no IR-radiation based on internal decomposition metabolism could be observed in wild boar carcasses at average ambient temperatures below 10 °C, and that this observation applied in spite of the presence of blowflies. Further details are also described in Probst et al. (2020). Under such circumstances, the IR-radiation of both the wild boar carcasses and the wild boar dummies was almost exclusively the result of external energy inputs. The masking effect was superior.
To increase the efficiency of thermal imaging techniques for detecting carcasses, based on our findings, we recommend that:
- During the vegetation period (average ambient temperature > 10 °C; April–October), airborne thermal imagers should mainly be used in open areas but preferably on overcast days. The best results can be achieved during nights following cloudy days.
- Outside of the vegetation period (average ambient temperatures 10 °C; November–March), probably only a short timeframe (before algor mortis is complete until the second day after death) can be used (Overview given in Table 4).
Based on these limitations, and keeping in mind that the simultaneous detection of living animals is also always a risk – especially during resting periods when animals are immobile – we recommend that an effective carcass detection regime should not only be based on the use of thermal imaging techniques, if such techniques are used at all. Considering that wild boar prefer dense vegetation, the use of specially trained dogs would be a more promising approach for the detection of wild boar carcasses during an ASF outbreak.
Acknowledgement
This study was only possible because of the help and support of many other persons. It was a pleasure for us to collaborate with the team of the Polizeihubschrauberstaffel Rheinland-Pfalz Winningen, headed by Jürgen Noll. We wish to thank Mike Brando (FAWF) and Ulrich Franke (wildlife monitoring by aerosense) for providing the technical equipment for ground measurements and supervision of data collection. Michael Otto helped us to find a fenced place for positioning the carcasses as well as the landowners Werner Dötsch and the Scherschlicht
family. We especially appreciated the uncomplicated cooperation with the Tierpark Daun for providing the study objects.
Ethical approval
The authors declare that no research interventions or experiments with animals or human beings, no clinical trials as well as no clinical research were conducted in the context of this study.
Conflict of interest
The authors declare that they have no conflicts of interest and no affiliations with or involvement in any organization or entity with any financial interest in the subject matter or materials discussed in this manuscript.
Funding
The authors declare that they had no financial support for this publication. We agree to make all data available upon reasonable request.
Authors contribution
UH: Project development, literature search and review, manuscript writing, manuscript editing.
MK: Field work and data processing. The summer part was topic of master thesis.
MS: Field work and data processing. The winter part was topic of master thesis.
FS: Supervision of master students. Checking manuscript.
Address for correspondence
Ulf Hohmann
Research Institute for Forest Ecology and Forestry
Rhineland-Palatinate
Hauptstr. 16
67705 Trippstadt, Germany
ulf.hohmann@wald-rlp.de
References
Amendt J, Rodner S, Schuch CP, Sprenger H, Weidlich L, Reckel F (2017): Helicopter thermal imaging for detecting insect infested cadavers. Sci Justice 57(5): 366–372.
Anderson GS (2000): Minimum and maximum development rates of some forensically important Calliphoridae (Diptera). J Forensic Sci 45(4): 824–832.
Bodnar SR, Ciotti J, Soroka J, Larsen RA, Sprague JE (2019): Drone-Assisted Thermal Imaging to Determine the Location of Unmarked Graves. J Forensic Identif 69(3): 378–392.
Butters O, Krosch MN, Roberts M, MacGregor D (2020): Application of forward-looking infrared (FLIR) imaging from an unmanned aerial platform in the search for decomposing remains. J Forensic Sci 66(1): 347–355.
Chenais E, Depner K, Guberti V, Dietze K, Viltrop A, Ståhl K (2019): Epidemiological considerations on African swine fever in Europe 2014–2018. Porcine Health Manage 5(1): 1–10.
Descalzi M (2019): Detection of Larval Aggregations Using a Drone Mounted Thermal Imaging Camera. Univ. of New Haven, USA, Master’s thesis.
DesMarais AM (2014): Detection of cadaveric remains by thermal imaging cameras. J Forensic Identif 64: 489–512.
Edelman GJ, Hoveling RJ, Roos M, van Leeuwen TG, Aalders MC (2013): Infrared imaging of the crime scene: possibilities and pitfalls. J Forensic Sci 58(5): 1156–1162.
Johnson AP, Mikac KM, Wallman JF (2013): Thermogenesis in decomposing carcasses. Forensic Sci Int 231(1-3): 271–277.
Kelly JA (2006): The influence of clothing, wrapping and physical trauma on carcass decomposition and arthropod succession in central South Africa. Bloemfontein, South Africa, University of the Free State, Diss.
Lee MJ, Voss SC, Franklin D, Dadour IR (2018): Preliminary investigation of aircraft mounted thermal imaging to locate decomposing remains via the heat produced by larval aggregations. Forensic Sci Int 289: 175–185.
McDaneld CP (2016): The effect of plastic tarps on the rate of human decomposition during the spring/summer in central Texas. San Marcos, Texas State University, USA, Diss.
Miteva A, Papanikolaou A, Gogin A, Boklund A, Bøtner A, Linden A, Viltrop A, Gortazar Schmidt C, Ivanciu C, Desmecht D, Korytarova D, Olsevskis E, Helyes G, Wozniakowski G, Thulke HH, Roberts H, Abrahantes JC, Stahl K, Depner K, Gonzalez Villeta LC, Spiridon M, Ostojic S, More S, Chesnoiu Vasile T, Grigaliuniene V, Guberti V, Wallo R (2020): EFSA (European Food Safety Authority) – Scientific report on the epidemiological analyses of African swine fever in the European Union (November 2018 to October 2019). EFSA J 18(1): 5996. https://doi.org/10.2903/j.efsa.2020.5996.
Morelle K, Jezek M, Licoppe A, Podgorski T (2019): Deathbed choice by ASF-infected wild boar can help find carcasses. Transbound Emerg Dis 66(5): 1821–1826.
Murray B, Anderson DT, Wescott DJ, Moorhead R, Anderson MF (2018): Survey and insights into unmanned aerial-vehicle-based detection and documentation of clandestine graves and human remains. Hum Biol 90(1): 45–61.
Sharanowski BJ, Walker EG, Anderson GS (2008): Insect succession and decomposition patterns on shaded and sunlit carrion in Saskatchewan in three different seasons. Forensic Sci Int 179: 219–240.
Probst C, Gethmann J, Hohmann U, Knoll B, Amendt J, Teifke JP, Conraths FJ (2020): Zersetzungsstadien bei Wildschweinkadavern und wie die Liegezeit geschätzt werden kann.
Amtstierärztlicher Dienst (ATD) 2020/II, ISSN 0945-3296: 85–94.
Zani L, Masiulis M, Bušauskas P, Dietze K, Pridotkas G, Globig A, Blome S, Mettenleiter T, Depner K, Karvelienė B (2020): African swine fever virus survival in buried wild boar carcasses. Transbound Emerg Dis 67: 2086–2092.











Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2020-46-Hohmann.pdf (0.94 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2020-46-Hohmann-Table1.pdf (0.05 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2020-46-Hohmann-Table2.pdf (0.05 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2020-46-Hohmann-Table3.pdf (0.05 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2020-46-Hohmann-Table4.pdf (0.05 MB) herunterladen möchten


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}