
SARS und COVID: Neuartige Krankheitsausbrüche zoonotischen Ursprungs aus Fledermäusen
Berliner und Münchener Tierärztliche Wochenschrift 134, 1-10
DOI: 10.2376/1439-0299-2020-29
© Schlütersche Fachmedien GmbH. 2021
Publiziert: 03/2021
Summary
SARS and COVID-19 are two severe human diseases with coronaviruses SARS-CoV-1 and SARS-CoV-2 as causative agents, respectively. Both viruses are taxonomically closely related betacoronaviruses originating from zoonotic spill-over events of their ancestors, which are representatives of a huge number of coronaviruses present in bat reservoirs. Although until the identification of SARS as a new human disease entity, coronaviruses generally were considered mild human pathogens, but had been recognized as major pathogens for some animal species.
SARS and COVID-19 outbreaks both emerged in China, probably from wet markets, facilitated by conditions that favor interspecies spill-over due to direct or indirect contacts between wildlife species and humans. Both pathogens have demonstrated their ability to not only infect wildlife species and humans but to occasionally spillover from humans to pet and farmed carnivore species.
Despite numerous similarities between SARS and COVID-19, there are major differences in their epidemiological characteristics. The case fatality rate of the SARS epidemic is estimated to be approximately tenfold higher than that of COVID-19. However, less than 800 people died from SARS, whereas about half a million people succumbed to COVID-19 worldwide in the first half year of the ongoing pandemic. The explosive spread of SARS-CoV-2 was at least in part facilitated by transmission without or before onset of symptoms, which hindered implementation of effective control measures.
To be better prepared for future epidemics and pandemics originating from zoonotic pathogens like SARS-CoV and SARS-CoV-2, preparing in „peacetime“ is crucial while profiting from state-of-the-art scientific knowledge and international collaboration in a One Health approach.
Zusammenfassung
SARS und COVID-19 sind zwei schwere Erkrankungen des Menschen, die durch Coronaviren (SARS-CoV-1 und SARS-CoV-2) ausgelöst werden. Beide Viren sind taxonomisch eng verwandte Betacoronaviren, die aus einem zoonotischen Spill-Over aus einigen der zahlreichen Fledermauscoronaviren hervorgingen.
Obwohl Coronaviren bis zur SARS-Epidemie im Allgemeinen als milde humane Pathogene angesehen wurden, sind Coronaviren bei einigen Tierarten als Erreger schwerwiegender Infektionskrankheiten bekannt.
SARS und COVID-19 haben ihren Ursprung wahrscheinlich auf chinesischen Tiermärkten, die durch den direkten und indirekten Kontakt zwischen Wildtieren und Menschen günstige Bedingungen für einen artübergreifenden Spill-Over bieten. Als Zoonosen haben beide Erreger ihre Fähigkeit bewiesen, nicht nur Wildtierarten und Menschen zu infizieren, sondern gelegentlich auch auf Haus- und Nutztiere überzuspringen. Besonders Katzen und Frettchen sind empfänglich für diese respiratorischen Krankheitserreger. Jedoch wurde eine Infektion von Besitzern durch ihre Haustiere bisher nicht dokumentiert. Deshalb scheinen Katzen und Hunde bisher keine signifikante Rolle in der Epidemiologie von COVID-19 beim Menschen zu spielen.
Neben ihren Gemeinsamkeiten gibt es trotzdem bedeutende Unterschiede in der Epidemiologie von SARS und COVID-19. Die Sterblichkeitsrate bei SARS ist schätzungsweise etwa zehnmal höher als bei COVID-19, obwohl im ersten Halbjahr der Pandemie um ein Vielfaches mehr Menschen an COVID-19 gestorben sind als während der SARS-Epidemie. Die explosive Ausbreitung von SARS-CoV-2 wurde zumindest teilweise durch die Übertragung durch infizierte Personen ohne klinisch relevante Symptome erleichtert, was die Einführung von Kontrollmaßnahmen ernsthaft erschwerte.
Um auf zukünftige Epidemien und Pandemien von zoonotischen Krankheitserregern besser vorbereitet zu sein, ist in „Friedenszeiten“ die Vorbereitung durch internationale und interdisziplinäre Zusammenarbeit im Sinne des One-Health-
Konzeptes von entscheidender Bedeutung.
Introduction
Severe acute respiratory syndrome (SARS) is a collective term for febrile respiratory disease entities of humans, caused by one of two recently emerged human coronaviruses. These two betacoronaviruses, SARS-CoV-1 and SARS-CoV-2, have caused or cause different disease entities in humans: SARS and coronavirus disease-19 (COVID-19), respectively.
There are similarities between the two disease entities and their causative agents. Both viruses use ACE-2 (Angiotensin-converting enzyme 2) as their cellular receptor and have their origin in bats, in which their most likely ancestors have been identified (ICTV 2020, Menachery et al. 2015).
The first reports of SARS as an apparently new contagious atypical pneumonia, often with fatal outcome, in the Guangdong Province in southern China date from the end of 2002. The start of the epidemic was most probably associated with human contacts with wet-market-traded carnivore species for human consumption. SARS verifiably affected over 8000 people and has cost 774 patients‘ lives (WHO 2003). It spread to 29 countries on three continents within a year until it was brought under control in 2003 (WHO 2003).
Top Job:
COVID-19 also emerged from China, with the first cases also suspected to be associated with zoonotic transmission at a wet market in Wuhan at the end of 2019. The disease was also characterized by severe pneumonia and it rapidly spread worldwide from the country of origin, sparking a true pandemic within a matter of weeks. It has affected more than 55 million people with more than a million deaths until mid of November 2020 (Figure 1; ECDC 2020).
SARS-associated Coronaviruses – Zoonotic Viruses with Pandemic Potential
Before 2002 human coronaviruses that were responsible for seasonal colds and relatively mild respiratory diseases predominantly in children and the elderly, got little attention from medical doctors and clinical virologists. However, coronaviruses generally have the ability to infect a broad spectrum of mammalian or avian species and show a predominant tropism for respiratory or gastro-intestinal organ systems. Until recently most of our knowledge about the pathogenesis of coronaviruses was gained from the detailed characterization of infections with animal coronaviruses like mouse hepatitis virus (Weiss and Leibowitz 2011).
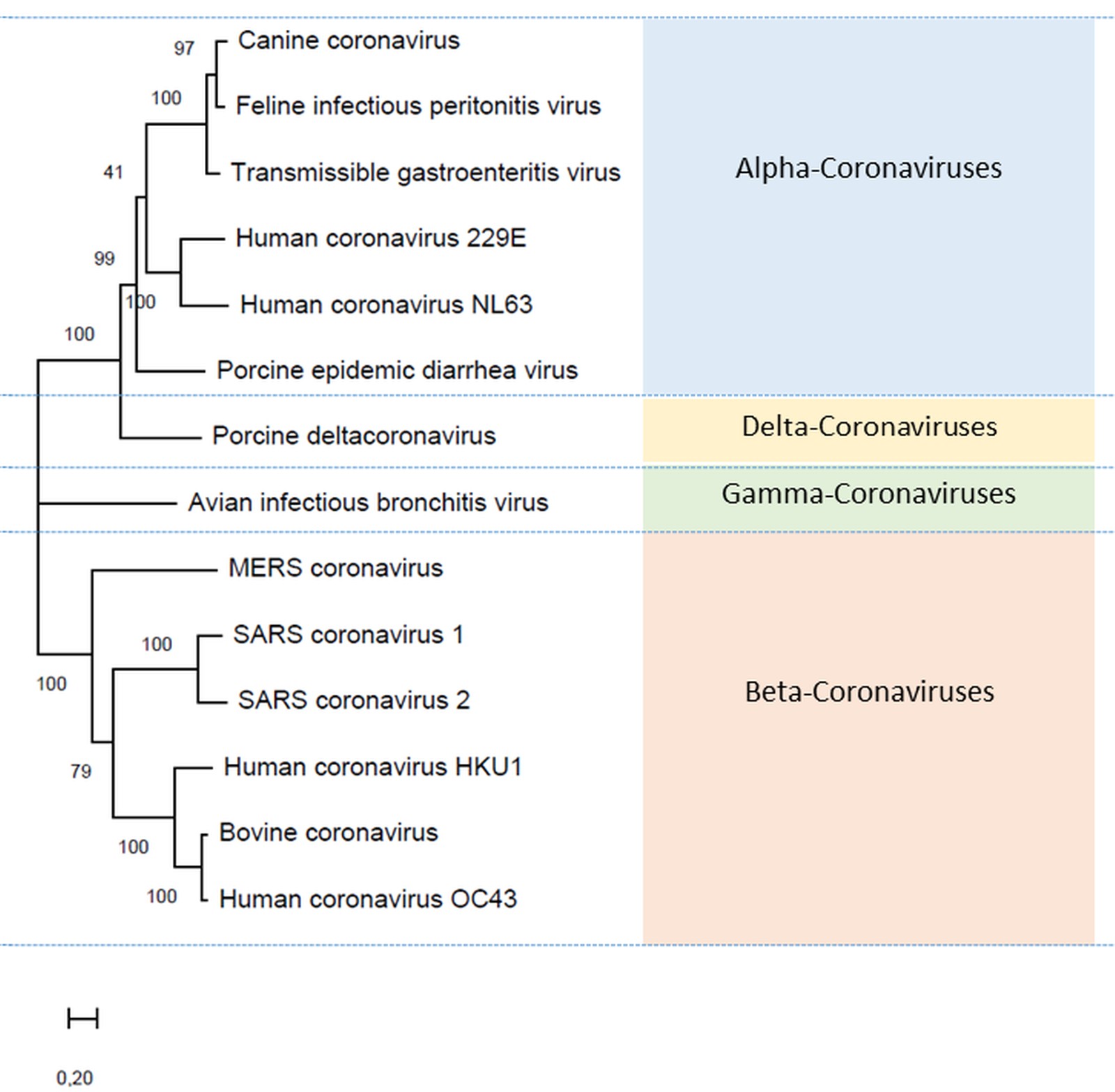
The family Coronaviridae of the order Nidovirales is divided in four subfamilies, of which members of the alpha- and the betacoronaviruses may cause more or less severe disease in humans and animals (ICTV 2020, Figure 2). The alphacoronaviruses harbour two members that may cause relatively mild upper respiratory disease in humans: HuCoV-229E and HuCoV-NL63. The betacoronaviruses also harbour two members that may cause equally mild upper respiratory disease in humans: HuCoV-OC43 and HuCoV-HKU1. In addition this subfamily harbours three members that have caused severe epidemics and a pandemic respectively: SARS-CoV-1, Middle East respiratory syndrome coronavirus (MERS-CoV) and SARS-CoV-2 (Gorbalenya et al. 2020). Coronaviruses have a single-stranded RNA genome with a positive polarity and an enveloped capsid.
Coronaviruses possess the largest genomes among RNA viruses comprising about 27.6–31 kb in size. Generally, the genome organization is defined by non-structural proteins (NSPs) which contain the main elements for virus replication like the RNA-dependend RNA polymerase. Whereas these NSPs are encoded by the first two-thirds of the genome, the last third contains mainly the genetic information for the four structural proteins, namely: spike protein (S), membrane protein (M), envelope protein (E) and the nucleocapsid protein (N) (Mortola and Roy 2004, Wang et al. 2017). Among these structural proteins the S protein is of special interest as it is an important determinant of host specificity and pathogenicity. The S protein forms the solar corona-like appearance around the viral particles under the electron microscope, hence the name coronavirus. A section of the S protein is responsible for receptor recognition (receptor binding domain – RBD) which defines the first point of interaction between the virion and the susceptible host cell. S protein domains also are involved in virus-cell fusion (Belouzard et al. 2012, Hulswit et al. 2016). For SARS-CoV-1 and SARS-CoV-2 it was demonstrated that both these betacoronaviruses use the cellular protease Angiotensin converting enzyme 2 (ACE2) as a cell entry receptor (Lan et al. 2020, Li et al. 2003). This mechanism of the S protein mediated cell entry may be an important target for antiviral and vaccine development.
Apart from their genome structure, coronaviruses are well known for their high mutation rate allowing for rapid adaptation to new host species, changing organ tropism and for antigenic plasticity. One major aspect leading to the progressive accumulation of mutations is the replicase, an RNA dependent-RNA polymerase lacking a sufficient proofreading function negatively influencing the nucleotide selectivity and fidelity (Neuman et al. 2014). Furthermore, co-infections with more than one coronavirus of different strains or viral species may lead to recombination events due to synthetizing a hybrid RNA combining sequences from the original viruses (Banner and Mc Lai 1991, Huang et al. 2016, Luytjes et al. 1988). This leaves coronaviruses equipped with the remarkable ability to change virulence, cell tropism, pathogenesis, and host range in a rapid and unpredictable way.
SARS – Two Emerging Respiratory Diseases
The four human coronaviruses HCoV-OC43, HCoV-NL63, HCoV-229E and HCoV-HKU1 are part of a heterologous group of viruses that circulate in the human population and may cause relatively mild upper respiratory disease or common colds, predominantly in children and older adults (Corman et al. 2018,
Su et al. 2016). Although until the identification of SARS as a new human disease entity, coronaviruses generally were considered mild human pathogens, they had been recognized as major pathogens for some animal species (Weiss and Leibowitz 2011). The more severe coronavirus infections in humans are caused by the three species of coronaviruses which were introduced into the human population within the last two decades: SARS-CoV-1, MERS-CoV and SARS-CoV-2. Here we only focus on the SARS related coronaviruses and their respective diseases.
The starting point of the first large coronavirus epidemic in the 21st century was defined by cases of atypical pneumonia in Fushan and neighbored cities in the South Chinese province Guangdong, where it emerged presumably in late 2002 (Zhao et al. 2003). The first major cluster was then recognized when the outbreak reached the capital city of Guangdong, Guangzhou where 226 patients were diagnosed until 8 February 2003 (Zhong et al. 2003). From there the virus spread quickly via neighboring Hong Kong along international air travel routes causing local clusters in different countries like Vietnam, Singapore and Canada (Hsu et al. 2003, Hwang et al. 2005, Lee et al. 2003, Riley et al. 2003, Tsang et al. 2003, Varia et al. 2003). Investigations of the causative agent led to the identification of a novel coronavirus as the etiological cause (Drosten et al. 2003, Fouchier et al. 2003, Ksiazek et al. 2003, Kuiken et al. 2003). The first complete genome sequences of the virus causing severe acute respiratory syndrome (SARS) were published in July 2003 and showed that the novel virus is only distantly related to the known human coronaviruses but showed the classic features of coronaviruses, like the genome organization (Marra et al. 2003, Rota et al. 2003). After the implementation of efficient control measures, including early case detection, extensive contact tracing and quarantine of contact people, the WHO announced that the global SARS outbreak was contained by 5 July 2003 (CDC 2020a, b, Hsu et al. 2003, Singh et al. 2003). Until July 2003 8098 SARS cases had been reported and of these 774 people had succumbed to the disease, marking a case fatality rate (CFR) of about 10% (CDC 2020a, b).
The SARS epidemic events remind us of the ongoing COVID-19 pandemic. Although SARS-CoV-2 emerged in Wuhan in the Chinese Province Hubei instead of Guangdong, the circumstances were rather similar.
Epidemiological investigations lead to the hypothesis that a wet market and/or wildlife animal traded there were at the epicenter of the outbreak at the end of 2019 (Lam et al. 2020). From Wuhan the virus spread rapidly to other provinces and countries despite efforts to contain the infection. The epidemic was ‘up-graded’ to a pandemic by WHO on 11 March 2020 after 118,000 confirmed cases in 114 countries and 4291 deaths had been reported (WHO 2020). Estimations of the case fatality rate pointed to different results influenced by the geographical area, local public health system, extent of testing in the population, methods of calculation and many more (Kim and Goel 2020, Rajgor et al. 2020). The initially estimated case fatality rates based on confirmed cases and associated deaths ranged between 0.7% in South Korea and 7.2% in Italy (Onder et al. 2020, Shim et al. 2020). To date we may conclude that although the overall case fatality rate of SARS was about ten times higher than that of COVID-2: the latter was associated with much higher morbidity and mortality due to its high transmissibility and worldwide spread.
Fatal Encounters – Interspecies Transmission of SARS-CoVs
The reconstruction of the zoonotic virus spill-over events from animals to humans has been a major challenge to virologists and epidemiologists alike. Until today there is no complete clarity about events that have led to the eventual incursion of SARS-CoVs into to the human population, from their postulated original bat hosts, most probably via intermediate hosts or host reservoirs.
Early epidemiological studies investigating the source of the SARS outbreak suggested a zoonotic transmission from an animal reservoir. The first indications came from the above average number of exotic animal traders from Guangzhou animal markets among the early detected SARS cases (Xu et al. 2004, Zhong et al. 2003). Additionally, serological surveillance showed a significantly higher prevalence of SARS-CoV IgG antibodies in animal traders in comparison to vegetable traders on Guangzhou wet markets (CDC 2003). Further investigations could confirm an infection of wildlife animals which were traded at the market. Subsequently, a virus was isolated from Himalayan palm civets (Paguma larvata) and raccoon dogs (Nyctereutes procyonoides) which displayed 99.8% sequence identity to SARS-CoV (Guan et al. 2003, Kan et al. 2005). The assumption that the animals got infected after their arrival at the animal market is supported by a study reporting the absence of SARS antibodies in civets from breeding farms (Tu et al. 2004) and wild civets (Poon et al. 2005). The fact that a widespread infection in wild or farmed civets is absent, together with the severity of disease the animals showed when infected (Wu et al. 2005) allowed to rule them out as a reservoir. Due to their high susceptibility these carnivores most likely functioned as an amplification factor and enabled adaptation of the virus to be sufficiently fit to infect humans. Although palm civets got into the focus as the intermediate host, other animal species traded at the wet markets were detected as SARS-CoV positive as well. Besides palm civets and raccoon dogs, surveillance of animals at live animal markets in January 2004, before animals of Guangdong Province markets were culled, identified four domestic cats, three red foxes and one Lesser rice rat, that were tested positive by RT-PCR (Wang et al. 2005).
A theory suggesting that civets could have got infected by a spill-over from bats which were also sold at the same animal market was verified by the discovery of a very closely related SARS-like coronavirus in the cave dwelling fruit bat (Rousettus leschenaultia) (Li et al. 2005) and of similar viruses in many other bat species (Lin et al. 2017).
Considering this information, it is assumed that the palm civet or the raccoon dog functioned as an intermediate host in the transmission chain from bats to humans. Still a simultaneous cross-over event from bats to humans and the suspected intermediate hosts or spill-over from humans to palm civets and raccoon dogs cannot be ruled out completely and remains subject of further speculation and study.
Similarly to the route via SARS-CoV found its way into the human population, it is speculated that SARS-CoV-2 benefited from infecting an intermediate host first (Liu et al. 2020). Although it is not clarified which animal served as the possible link between bats and humans, it has been postulated that the Malayan pangolin (Manis javanica) has played a role in this development as coronavirus spike protein sequences that show high similarity to SARS-CoV-2 spike protein have been identified in this species (Jaimes et al. 2020, Lam et al. 2020). However, later studies showed that it is unlikely that indeed the pangolin was the intermediate host (Latinne et al. 2020). The lack of detailed knowledge about the recent evolutionary routes and how the virus entered the human population gives space for many speculations and rumors. Theories that SARS-CoV-2 was artificially constructed or manipulated in the laboratory can be ruled out as the nucleotide sequence of SARS-CoV-2 shows mutations which are not seen in previously known coronavirus sequences and are most likely the result of natural selection and adaptation to the human host (Andersen et al. 2020). Also the furin-cleavage site in the spike protein, which is critical for the enhanced entry ability of SARS-CoV-2 into human cells is not an indication for human engineering, but seen in other bat coronaviruses (Zhou et al. 2020).
The fact that there is a Biosafety level 4 laboratory in Wuhan, where the virus was first detected, with scientists work on bat coronaviruses can be an unlucky coincidence. So far there is no evidence that it escaped from laboratory but it is hard to proof this theory wrong. As long as questions about the transmission route from bats into the human population remain unanswered, speculations and misinformation will be part of the debate.
Transmission between People and their Pets
The zoonotic origins of the SARS and COVID-19 outbreaks quickly raised public attention and discussions about the possibility of a COVID-19 transmission between humans and animals in close contact with them. The knowledge about the animal-to-human spread of these viruses had an impact on the public perception of domestic animals especially in China (Yin et al. 2020).
SARS
For SARS-CoV the concerns about possible infection chains involving domestic animals arose with epidemiological reports about the role of intermediate hosts as an amplifying factor and virus spreader. Investigations documented that, besides palm civets different carnivorous species like raccoon dogs and foxes and cats as well as rats in Guangdong Province tested positive by RT-PCR (Wang et al. 2005). Surveillance in domestic pigs could identify two SARS-CoV positive individuals which were probably infected by virus-contaminated feed rather than human-to-swine transmission (Chen et al. 2005). Extensive surveillance in livestock did not detect other infected species (Wang et al. 2005). Besides that data were published describing experimental infection experiments of laboratory monkeys (Fouchier et al. 2003, McAuliffe et al. 2004, Qin et al. 2005), ferrets and cats (Martina et al. 2003), mice (Glass et al. 2004, Wentworth et al. 2004), pigs and chicken (Weingartl et al. 2004), guinea pigs (Liang et al. 2005) and Syrian Gold Hamsters (Roberts et al. 2005). When evaluating the risk of a transmission between companion animals, it is worth mentioning that experimentally infected cats were able to infect naïve contact cats (Martina et al. 2003).
COVID-19
Soon after the beginning of the pandemic COVID-19 outbreak in early 2020 discussions about its origin in wildlife animals had a considerable impact on the relation of people to domestic animals and wildlife. One consequence was, that the fear of an additional spillover from coronavirus-harboring bats led to recommendations for mass killings of bat colonies, putting many endangered species at increased risk (Wang Hongxin et al. 2020, Zhao 2020). In the meantime more data about infections of domestic animals was obtained. Cases of SARS-CoV-2 positive companion-, farmed-, and zoo animals indicated that these animals may indeed be infected.
Among the companion animals, cats appear to be the pet species that is most susceptible to natural infection. Sporadic cases of SARS-CoV-2 positive cats have been diagnosed by RT-PCR in various countries (AFCD 2020, ScienceNews 2020, Sit et al. 2020). Investigating the circumstances lead to the assumption that these animals became most likely infected by their COVID-19 infected owners (Sit et al. 2020). Not surprisingly big cats species also turned out to be susceptible to infection, as a tiger in a U.S. zoological collection got infected, most probably by an animal care taker and displayed symptoms of respiratory disease (Wang Leyi et al. 2020). In addition to these isolated cases, data from experimental infections of cats indicate that cats are susceptible to a productive SARS-CoV-2 infection which is characterized by a prolonged period of oral and nasal virus shedding, although it is in most cases not associated with overt clinical signs of disease (Bosco-Lauth et al. 2020). In the same experimental setting, a transmission between SARS-CoV-2 inoculated and naïve contact cats could be documented. Furthermore several reports have documented that dogs may sporadically contract SARS-CoV-2 infection without overt clinical symptoms and displaying low virus loads (AFCD 2020, Goumenou et al. 2020). Data from experimentally infected dogs support the findings from infected pet dogs. After inoculation, viral RNA could be detected in rectal swab samples from these animals, although no replicating virus could be isolated (Shi et al. 2020). Similarly, serological surveillance among different livestock animals in China indicate that these animals play no role as intermediate hosts and are most likely not susceptible (Deng et al. 2020).
The feature of the pandemic to act as a reverse zoonosis and infect animals as the consequence of a spillover from humans was demonstrated in June 2020 when ten thousands of minks on Dutch mink farms had to be culled. This happened due to rapid spread among these animals after a probable introduction from COVID-19 diseased farm workers (Enserink 2020, Molenaar et al. 2020, Oreshkova et al. 2020). It was also shown that mink-to-human infection had occurred (LNV 2020). The high susceptibility of minks (Neovison vison) is not surprising as minks and ferrets are closely related members of the family Mustelidae. Among the susceptible animals, ferrets and Syrian golden hamsters (Mesocricetus auratus) have been shown to be suitable laboratory animal models for studying the pathogenesis of COVID-19 and its intervention strategies (Kim et al. 2020, Richard et al. 2020, Sia et al. 2020). To monitor the occurrence of this virus in pets, COVID-19 infections in pets are reportable to the authorities in Germany (BReg 328/20 2020).
Although SARS-CoV-1 and SARS-CoV-2 infections of domestic animals seem to have no significant role in the epidemiology of COVID-19 in humans, and with the exception of a documented mink-to-human infection, there is no evidence for animal-to-human transmission, these reports point to the assumption that infections of animals living in close contact with humans can occur. Even when considering that SARS-CoV-2 may be transmitted across species barriers, SARS-CoV-2 infections of pets by infected owners appear to be rare events (Leroy et al. 2020).
Prevention and Intervention Strategies – A One Health Approach
Both the emergence of SARS and COVID-19 share notable similarities with other emerging viral diseases that originate from zoonotic events. A better understanding of the factors that lead to zoonotic and subsequent human-to-human transmission, may help to develop more effective intervention strategies for future epidemics and pandemics. As the associated morbidity and mortality, challenge society at multiple levels worldwide, international, scientific collaboration among all related disciplines, as well as communication among all stakeholders using a One Health approach appears to be the most promising strategy.
The likelihood for virus spill-over events into the human population depends on direct or indirect contacts between people and the reservoir hosts. The opportunities for such spill-over events have significantly increased in the past decades due to our globalizing society in the Anthropocene era, which is characterized by encroachment of wildlife habitats, intensification of livestock rearing as well as the increasing spread of arthropod-borne pathogens due to climate change (Jones et al. 2008, Reperant et al. 2012). Zoonotic virus transmissions that have led to major epidemics or pandemics like AIDS, Influenza, SARS, COVID-19, Nipah or Ebola imply that direct or indirect contacts to wildlife, especially non-human primates, birds and bats are a major risk factor (Han et al. 2015). Trading and consumption of wildlife and deforestation are examples of driving factors which unfortunately are hard to control (Cantlay et al. 2017).
Another important aspect of adequately dealing with major epidemics or pandemics is the implementation of early warning systems for disease outbreaks in human and animals alike. These surveillance and reporting systems should be based upon state-of-the-art syndrome reporting and laboratory diagnosis. This requires collaboration between national and international organizations like the World Health Organization (WHO), the Organization for Animal Health (OIE) and Food and Agricultural Organization (FAO). The implementation of surveillance and diagnostic capacities allowing early identification of an emerging pathogen is crucial to slow down its spread, saving essential time for further action. Subsequently non-pharmaceutical and pharmaceutical intervention (NPI and PI) strategies should then be applied without losing time. To respond as fast as possible, it is essential to make the necessary preparations in “peacetime”. In following this strategy, for SARS it was possible indeed to control the epidemic with basic public health measures (Lipsitch et al. 2003).
During the ongoing COVID-19 pandemic numerous NPIs like widespread testing, contact tracing and isolation, as well as avoidance of superspreading events, have proven to be crucial, but were not sufficient to stop the pandemic spread of SARS-CoV-2 (Choi 2020, Cousins 2020, Steinbrook 2020). Other key factors are related to the resilience of public health systems like hospital and ICU (intensive care unit) capacity (Grasselli et al. 2020). Not only is an adequate number of intensive care beds needed, but also sufficient protective equipment as nosocomial transmission is putting essential health care workers at risk (Gamage et al. 2005, Lancet 2020).
To understand which factors might contribute to potential outbreak situations, the role of wildlife and exotic animal trade for collections or for consumption should not be underestimated and needs further investigation (Chomel et al. 2007, Karesh et al. 2005).
Another important step for better preparedness against emerging virus outbreaks is gathering more knowledge about the global virome, by investing in state-of-the-art pathogen discovery and characterization platforms (Greninger 2018, Sridhar et al. 2015). The tools for rapid identification and characterization of viruses in their natural reservoirs and in newly invaded hosts is of crucial importance.
In addition to these measures, for the best possible preparedness, rapid development of broadly protective antiviral therapeutics and vaccines should be a priority. To complement the rapid development antiviral therapeutics, quick repurposing of already known antivirals can be a helpful additional strategy. To rapidly obtain safe and effective antiviral therapeutics and vaccines, investment in state-of-the-art product development pipelines and vaccine development platforms in “peacetime” will contribute to the timely availability of sufficient antiviral and vaccine candidates which can be rapidly adapted to a newly emerging virus (Afrough et al. 2019).
Conclusion
In the past two decades, the world has been confronted with three newly identified human coronavirus diseases of which the causative agents had spilled-over from animal reservoirs. Two of these, SARS and COVID-2, share many features, but differ in a higher case fatality rate for SARS-CoV-1 and in a higher transmissibility for SARS-CoV-2. Consequently, SARS-CoV-2 proved to be more difficult to contain and caused a still ongoing pandemic for which today no specific antiviral treatment or vaccine are available.
It is a matter of time until yet another virus from its wildlife reservoir will be transmitted to humans, and cause yet another epidemic or even pandemic. Definitely, coronaviruses are among the lead candidates for such events.
To be better prepared for future epidemics and pandemics originating from the animal world it is crucial to learn from the ongoing COVID-19 and preceding pandemics originating from zoonotic pathogens. Preparing in “peacetime“ is crucial, profiting from state-of-the-art scientific knowledge and international collaboration in a One Health approach.
Ethical approval
Not applicable.
Conflict of interest
The authors declare that there is no conflict of interest.
Funding
This work was supported by Deutsche Forschungsgemeinschaft (DFG; German Research Foundation) -398066876/GRK 2485/1. The authors hereby agree to provide the details of such funding upon a reasonable request.
Authors contribution
Manuscript writing: FK.
Discussing and final editing: AO.
Address for correspondence
Dr. med. vet. Franziska Kaiser
Research Center for Emerging Infections and Zoonoses
Bünteweg 17
30559 Hannover
franziska.kaiser@tiho-hannover.de
References
Afrough B, Dowall S, Hewson R (2019): Emerging viruses and current strategies for vaccine intervention. J Clin Exp Immunol 196(2): 157–166.
Agriculture, Fisheries and Conservation Department of Hong Kong (AFCD) (2020): Pet cat tests positive for COVID-19 virus. https://www.info.gov.hk/gia/general/202003/31/P2020033100717.htm (accessed 06.08.2020).
Andersen KG, Rambaut A, Lipkin WI, Holmes EC, Garry RF (2020): The proximal origin of SARS-CoV-2. Nat Med 26(4): 450–452.
Banner LR, Mc Lai M (1991): Random nature of coronavirus RNA recombination in the absence of selection pressure. Virology 185(1): 441–445.
Belouzard S, Millet JK, Licitra BN, Whittaker GR (2012): Mechanisms of coronavirus cell entry mediated by the viral spike protein. Viruses 4(6): 1011–1033.
Bosco-Lauth AM, Hartwig AE, Porter SM, Gordy PW, Nehring M, Byas AD, VandeWoude S, Ragan IK, Maison RM, Bowen RA (2020): Pathogenesis, transmission and response to re-exposure of SARS-CoV-2 in domestic cats. BioRxiv: 2020.05.28.120998.
BReg 328/20 (2020): Dritte Verordnung zur Änderung der Verordnung über meldepflichtige Tierkrankheiten. Drucksache 328/20. https://www.bundesrat.de/SharedDocs/drucksachen/2020/0301-0400/328-20.p… (accessed 01.02.2021).
Cantlay JC, Ingram DJ, Meredith AL (2017): A Review of Zoonotic Infection Risks Associated with the Wild Meat Trade in Malaysia. EcoHealth 14(2): 361–388.
Center for Disease Control and Prevention (CDC) (2003): Prevalence of IgG antibody to SARS-associated coronavirus in animal traders-Guangdong Province, China, 2003. MMWR 52(41): 986–987.
Center for Disease Control and Prevention (CDC) (2020a): https://www.cdc.gov/about/history/sars/timeline.htm (accessed 07.08.2021).
Centers for Disease Control and Prevention (CDC) (2020b): https://www.cdc.gov/sars/about/faq.html (accessed 07.08.2021).
Chen W, Yan M, Yang L, Ding B, He B, Wang Y, Liu X, Liu C, Zhu H, You B, Huang S, Zhang J, Mu F, Xiang Z, Feng X, Wen J, Fang J, Yu J, Yang H, Wang J (2005): SARS-associated coronavirus transmitted from human to pig. Emerg Infect Dis 11(3): 446–448.
Choi JY (2020): COVID-19 in South Korea. Postgrad Med J 96(1137): 399–402.
Chomel BB, Belotto A, Meslin F-X (2007): Wildlife, exotic pets, and emerging zoonoses. Emerg Infect Dis 13(1): 6–11.
Corman VM, Muth D, Niemeyer D, Drosten C (2018): Hosts and Sources of Endemic Human Coronaviruses. Adv Virus Res 100: 163–188.
Cousins S (2020): New Zealand eliminates COVID-19. Lancet 395(10235): 1474.
Deng J, Jin Y, Liu Y, Sun J, Hao L, Bai J, Huang T, Lin D, Jin Y, Tian K (2020): Serological survey of SARS-CoV-2 for experimental, domestic, companion and wild animals excludes intermediate hosts of 35 different species of animals. Transbound Emerg Dis 67(4): 1745–1749.
Drosten C, Günther S, Preiser W, van der Werf S, Brodt H-R, Becker S, Rabenau H, Panning M, Kolesnikova L, Fouchier RAM, Berger A, Burguière A-M, Cinatl J, Eickmann M, Escriou N, Grywna K, Kramme S, Manuguerra J-C, Müller S, Rickerts V, Stürmer M, Vieth S, Klenk HD, Osterhaus ADME, Schmitz H, Doerr HW (2003): Identification of a novel coronavirus in patients with severe acute respiratory syndrome. N Engl J Med 348(20): 1967–1976.
Enserink M (2020): Coronavirus rips through Dutch mink farms, triggering culls. Science 368(6496): 1169.
European Centre for Diseasae Prevention and Control (ECDC) (2020): COVID-19 situation update worldwide. https://www.ecdc.europa.eu/en/geographical-distribution-2019-ncov-cases (accessed 07.08.2020).
Fouchier RAM, Kuiken T, Schutten M, Van Amerongen G, Van Doornum GJJ, Van Den Hoogen BG, Peiris M, Lim W, Stöhr K, Osterhaus ADME (2003): Aetiology: Koch’s postulates fulfilled for SARS virus. Nature 423(6937): 240.
Gamage B, Moore D, Copes R, Yassi A, Bryce E (2005): Protecting health care workers from SARS and other respiratory pathogens: a review of the infection control literature. Am J Infect Control 33(2): 114–121.
Glass WG, Subbarao K, Murphy B, Murphy PM (2004): Mechanisms of host defense following severe acute respiratory syndrome-coronavirus (SARS-CoV) pulmonary infection of mice. J Immunol 173: 4030–4039.
Gorbalenya AE, Baker SC, Baric RS, de Groot RJ, Drosten C, Gulyaeva AA, Haagmans BL, Lauber C, Leontovich AM, Neuman BW, Penzar D, Perlman S, Poon LLM, Samborskiy DV, Sidorov IA, Sola I, Ziebuhr J (2020): The species Severe acute respiratory syndrome-related coronavirus: classifying 2019-nCoV and naming it SARS-CoV-2. Nat Microbiol 5(4): 536–544.
Goumenou M, Spandidos AD, Tsatsakis A (2020): Possibility of transmission through dogs being a contributing factor to the extreme Covid‑19 outbreak in North Italy. Mol Med Rep 21(6): 2293–2295.
Grasselli G, Pesenti A, Cecconi M (2020): Critical Care Utilization for the COVID-19 Outbreak in Lombardy, Italy: Early Experience and Forecast During an Emergency Response. J Am Med Assoc 323(16): 1545–1546.
Greninger AL (2018): A decade of RNA virus metagenomics is (not) enough. Virus Res 244: 218–229.
Guan Y, Zheng BJ, He YQ, Liu XL, Zhuang ZX, Cheung CL, Luo SW, Li PH, Zhang LJ, Guan YJ, Butt KM, Wong KL, Chan KW, Lim W, Shortridge KF, Yuen KY, Peiris JSM, Poon LLM (2003): Isolation and Characterization of Viruses Related to the SARS Coronavirus from Animals in Southern China. Science 302(5643): 276–278.
Han H-J, Wen H, Zhou C-M, Chen F-F, Luo L-M, Liu J, Yu X-J (2015): Bats as reservoirs of severe emerging infectious diseases. Virus Res 205: 1–6.
Hsu L-Y, Lee C-C, Green JA, Ang B, Paton NI, Lee L, Villacian JS, Lim P-L, Earnest A, Leo Y-S (2003): Severe acute respiratory syndrome (SARS) in Singapore: clinical features of index patient and initial contacts. Emerg Infect Dis 9(6): 713–717.
Huang C, Liu WJ, Xu W, Jin T, Zhao Y, Song J, Shi Y, Ji W, Jia H, Zhou Y, Wen H, Zhao H, Liu H, Li H, Wang Q, Wu Y, Wang L, Liu D, Liu G, Yu H, Holmes EC, Lu L, Gao GF (2016): A Bat-Derived Putative Cross-Family Recombinant Coronavirus with a Reovirus Gene. PLoS Pathog 12(9): e1005883.
Hulswit RJG, de Haan CAM, Bosch B-J (2016): Coronavirus Spike Protein and Tropism Changes. Adv Virus Res 96: 29–57.
Hwang DM, Chamberlain DW, Poutanen SM, Low DE, Asa SL, Butany J (2005): Pulmonary pathology of severe acute respiratory syndrome in Toronto. Mod Pathol 18(1): 1–10.
International Committee on Taxonomy of Viruses (ICTV) (2020): The species Severe acute respiratory syndrome-related coronavirus: classifying 2019-nCoV and naming it SARS-CoV-2. Nat Microbiol 5(4): 536–544.
Jaimes JA, André NM, Chappie JS, Millet JK, Whittaker GR (2020): Phylogenetic Analysis and Structural Modeling of SARS-CoV-2 Spike Protein Reveals an Evolutionary Distinct and Proteolytically Sensitive Activation Loop. J Mod Biol 432(10): 3309–3325.
Jones KE, Patel NG, Levy MA, Storeygard A, Balk D, Gittleman JL, Daszak P (2008): Global trends in emerging infectious diseases. Nature 451(7181): 990–993.
Kan B, Wang M, Jing H, Xu H, Jiang X, Yan M, Liang W, Zheng H, Wan K, Liu Q, Cui B, Xu Y, Zhang E, Wang H, Ye J, Li G, Li M, Cui Z, Qi X, Chen K, Du L, Gao K, Zhao YT, Zou XZ, Feng YJ, Gao YF, Hai R, Yu D, Guan Y, Xu J (2005): Molecular evolution analysis and geographic investigation of severe acute respiratory syndrome coronavirus-like virus in palm civets at an animal market and on farms. J Virol 79(18): 11892–11900.
Karesh WB, Cook RA, Bennett EL, Newcomb J (2005): Wildlife trade and global disease emergence. Emerg Infect Dis 11(7): 1000–1002.
Kim DD, Goel A (2020): Estimating case fatality rates of COVID-19. Lancet Infect Dis 20(7): 773–774.
Kim YI, Kim S-G, Kim S-M, Kim E-H, Park S-J, Yu K-M, Chang J-H, Kim EJ, Lee S, Casel MAB, Um J, Song M-S, Jeong HW, Lai VD, Kim Y, Chin BS, Park J-S, Chung K-H, Foo S-S, Poo H, Mo IP, Lee OJ, Webby RJ, Jung JU, Choi YK (2020): Infection and Rapid Transmission of SARS-CoV-2 in Ferrets. Cell Host Microbe 27(5): 704–709.e2.
Ksiazek TG, Erdman D, Goldsmith CS, Zaki SR, Peret T, Emery S, Tong S, Urbani C, Comer JA, Lim W, Rollin PE, Dowell SF, Ling A-E, Humphrey CD, Shieh W-J, Guarner J, Paddock CD, Rota P, Fields B, DeRisi J, Yang JY, Cox N, Hughes JM, LeDuc JW, Bellini WJ, Anderson LJ; SARS Working Group (2003): A novel coronavirus associated with severe acute respiratory syndrome. N Engl J Med 348(20): 1953–1966.
Kuiken T, Fouchier RAM, Schutten M, Rimmelzwaan GF, van Amerongen G, van Riel D, Laman JD, de Jong T, van Doornum G, Lim W, Ling AE, Chan PKS, Tam JS, Zambon MC, Gopal R, Drosten C, van der Werf S, Escriou N, Manuguerra J-C, Stöhr K, Peiris JSM, Osterhaus ADME (2003): Newly discovered coronavirus as the primary cause of severe acute respiratory syndrome. Lancet 362(9380): 263–270.
Lam TT-Y, Jia N, Zhang Y-W, Shum MH-H, Jiang J-F, Zhu H-C, Tong Y-G, Shi Y-X, Ni X-B, Liao Y-S, Li W-J, Jiang B-G, Wei W, Yuan T-T, Zheng K, Cui X-M, Li J, Pei G-Q, Qiang X, Cheung WYM, Li LF, Sun FF, Qin S, Huang JC, Leung GM, Holmes EC, Hu YL, Guan Y, Cao W-C (2020): Identifying SARS-CoV-2-related coronaviruses in Malayan pangolins. Nature 583(7815): 282–285.
Lan J, Ge J, Yu J, Shan S, Zhou H, Fan S, Zhang Q, Shi X, Wang Q, Zhang L, Wang X (2020): Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 581(7807): 215–220.
Lancet (2020): COVID-19: protecting health-care workers. Lancet 395(10228): 922.
Latinne A, Hu B, Olival K, Zhu G, Zhang L, Li H, Chmura A, Field H, Zambrana-Torrelio C, Epstein J, Li B, Zhang W, Wang LF, Shi ZL, Daszak P (2020): Origin and cross-species transmission of bat coronaviruses in China. Nat Commun 11: 4235.
Lee N, Hui D, Wu A, Chan P, Cameron P, Joynt GM, Ahuja A, Yung MY, Leung CB, To KF, Lui SF, Szeto CC, Chung S, Sung JJY (2003): A major outbreak of severe acute respiratory syndrome in Hong Kong. N Engl J Med 348(20): 1986–1994.
Leroy EM, Ar Gouilh M, Brugère-Picoux J (2020): The risk of SARS-CoV-2 transmission to pets and other wild and domestic animals strongly mandates a one-health strategy to control the COVID-19 pandemic. One Health 10: 100133.
Li W, Moore MJ, Vasilieva N, Sui J, Wong SK, Berne MA, Somasundaran M, Sullivan JL, Luzuriaga K, Greenough TC, Choe H, Farzan M (2003): Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 426(6965): 450–454.
Li W, Shi Z, Yu M, Ren W, Smith C, Epstein J, Wang H, Crameri G, Hu Z, Zhang H, Zhang J, McEachern J, Field H, Daszak P, Eaton BT, Zhang S, Wang L-F (2005): Bats Are Natural Reservoirs of SARS-Like Coronaviruses. Science 310(5748): 676–679.
Liang L, He C, Lei M, Li S, Hao Y, Zhu H, Duan Q (2005): Pathology of guinea pigs experimentally infected with a novel reovirus and coronavirus isolated from SARS patients. DNA Cell Biol 24(8): 485–490.
Lin X-D, Wang W, Hao Z-Y, Wang Z-X, Guo W-P, Guan X-Q, Wang M-R, Wang H-W, Zhou R-H, Li M-H, Tang G-P, Wu J, Holmes EC, Zhang Y-Z (2017): Extensive diversity of coronaviruses in bats from China. Virology 507: 1–10.
Lipsitch M, Cohen T, Cooper B, Robins J, Ma S, James L, Gopalakrishna G, Chew S, Tan C, Samore M, Fisman D, Murray M (2003): Transmission Dynamics and Control of Severe Acute Respiratory Syndrome. Science 300(5627): 1966–1970.
Liu Z, Xiao X, Wei X, Li J, Yang J, Tan H, Zhu J, Zhang Q, Wu J, Liu L (2020): Composition and divergence of coronavirus spike proteins and host ACE2 receptors predict potential intermediate hosts of SARS-CoV-2. J Med Virol 92(6): 595–601.
LNV – Ministerie van Landbouw, Natuur en Voedselkwaliteit (2020): New results from research into COVID-19 on mink farms. https://www.government.nl/latest/news/2020/05/19/new-results-from-resea… (accessed 07.08.2020).
Luytjes W, Bredenbeek PJ, Noten AF, Horzinek MC, Spaan WJ (1988): Sequence of mouse hepatitis virus A59 mRNA 2: indications for RNA recombination between coronaviruses and influenza C virus. Virology 166(2): 415–422.
Marra MA, Jones SJM, Astell CR, Holt RA, Brooks-Wilson A, Butterfield YSN, Khattra J, Asano JK, Barber SA, Chan SY, Cloutier A, Coughlin SM, Freeman D, Girn N, Griffith OL, Leach SR, Mayo M, McDonald H, Montgomery SB, Pandoh PK, Petrescu AS, Robertson AG, Schein JE, Siddiqui A, Smailus DE, Stott JM, Yank GS, Plummer F, Andonov A, Artsob H, Bastien N, Bernard K, Booth TF, Bowness D, Czub M, Drebot M, Fernando L, Flick R, Garbutt M, Gray M, Grolla A, Jones S, Feldmann H, Meyers A, Kabani A, Li Y, Normand S, Stroher U, Tipples GA, Tyler S, Vogrig R, Ward D, Watson B, Brunham RC, Krajden M, Petric M, Skowronski DM, Upton C, Roper RL (2003): The Genome sequence of the SARS-associated coronavirus. Science 300(5624): 1399–1404.
Martina BEE, Haagmans BL, Kuiken T, Fouchier RAM, Rimmelzwaan GF, Van Amerongen G, Peiris JSM, Lim W, Osterhaus ADME (2003): SARS virus infection of cats and ferrets. Nature 425(6961): 915.
McAuliffe J, Vogel L, Roberts A, Fahle G, Fischer S, Shieh W-J, Butler E, Zaki S, St Claire M, Murphy B, Subbarao K (2004): Replication of SARS coronavirus administered into the respiratory tract of African Green, rhesus and cynomolgus monkeys. Virology 330(1): 8–15.
Menachery VD, Yount BLJ, Debbink K, Agnihothram S, Gralinski LE, Plante JA, Graham RL, Scobey T, Ge X-Y, Donaldson EF, Randell SH, Lanzavecchia A, Marasco WA, Shi Z-L, Baric RS (2015): A SARS-like cluster of circulating bat coronaviruses shows potential for human emergence. Nat Med 21(12): 1508–1513.
Molenaar RJ, Vreman S, Hakze-van der Honing RW, Zwart R, de Rond J, Weesendorp E, Smit LAM, Koopmans M, Bouwstra R, Stegeman A, van der Poel WHM (2020): Clinical and Pathological Findings in SARS-CoV-2 Disease Outbreaks in Farmed Mink (Neovison vison).Vet Pathol 57: 653–657.
Mortola E, Roy P (2004): Efficient assembly and release of SARS coronavirus-like particles by a heterologous expression system. FEBS Lett 576(1–2): 174–178.
Neuman BW, Chamberlain P, Bowden F, Joseph J (2014): Atlas of coronavirus replicase structure. Virus Res 194: 49–66.
Onder G, Rezza G, Brusaferro S (2020): Case-Fatality Rate and Characteristics of Patients Dying in Relation to COVID-19 in Italy. J Am Med Assoc 323(16): 1619.
Oreshkova N, Molenaar RJ, Vreman S, Harders F, Oude Munnink BB, Hakze-van der Honing RW, Gerhards N, Tolsma P, Bouwstra R, Sikkema RS, Tacken MG, de Rooij MM, Weesendorp E, Engelsma MY, Bruschke CJ, Smit LA, Koopmans M, van der Poel WH, Stegeman A (2020): SARS-CoV-2 infection in farmed minks, the Netherlands, April and May 2020. Euro Surveill 25: 2001005.
Poon LL, Chu DK, Chan KH, Wong OK, Ellis TM, Leung YH, Lau SK, Woo PC, Suen KY, Yuen KY, Guan Y, Peiris JS (2005): Identification of a novel coronavirus in bats. J Virol 79: 2001–2009.
Qin C, Wang J, Wei Q, She M, Marasco WA, Jiang H, Tu X, Zhu H, Ren L, Gao H, Guo L, Huang L, Yang R, Cong Z, Guo L, Wang Y, Liu Y, Sun Y, Duan S, Qu J, Chen L, Tong W, Ruan L, Liu P, Zhang H, Zhang J, Zhang H, Liu D, Liu Q, Hong T, He W (2005): An animal model of SARS produced by infection of Macaca mulatta with SARS coronavirus. J Pathol 206(3): 251–259.
Rajgor DD, Lee MH, Archuleta S, Bagdasarian N, Quek SC (2020): The many estimates of the COVID-19 case fatality rate. Lancet Infect Dis 20(7): 776–777.
Reperant LA, Kuiken T, Osterhaus ADME (2012): Adaptive pathways of zoonotic influenza viruses: from exposure to establishment in humans. Vaccine 30(30): 4419–4434.
Richard M, Kok A, de Meulder D, Bestebroer TM, Lamers MM, Okba NMA, Fentener van Vlissingen M, Rockx B, Haagmans BL, Koopmans MPG, Fouchier RAM, Herfst S (2020): SARS-CoV-2 is transmitted via contact and via the air between ferrets. Nat Commun 11(1): 3496.
Riley S, Fraser C, Donnelly CA, Ghani AC, Abu-Raddad LJ, Hedley AJ, Leung GM, Ho L-M, Lam T-H, Thach TQ, Chau P, Chan K-P, Lo S-V, Leung P-Y, Tsang T, Ho W, Lee K-H, Lau EMC, Ferguson NM, Anderson RM (2003): Transmission Dynamics of the Etiological Agent of SARS in Hong Kong: Impact of Public Health Interventions. Science 300(5627): 1961–1966.
Roberts A, Vogel L, Guarner J, Hayes N, Murphy B, Zaki S, Subbarao K (2005): Severe Acute Respiratory Syndrome Coronavirus Infection of Golden Syrian Hamsters. J Virol 79(1): 503 LP–511.
Rota PA, Oberste MS, Monroe SS, Nix WA, Campagnoli R, Icenogle JP, Peñaranda S, Bankamp B, Maher K, Chen M-H, Tong S, Tamin A, Lowe L, Frace M, DeRisi JL, Chen Q, Wang D, Erdman DD, Peret TCT, Burns C, Ksiazek TG, Rollin PE, Sanchez A, Liffick S, Holloway B, Limor J, McCaustland K, Olsen-Rasmussen M, Fouchier R, Günther S, Osterhaus ADME, Drosten C, Pallansch MA, Anderson LJ, Bellini WJ (2003): Characterization of a novel coronavirus associated with severe acute respiratory syndrome. Science 300(5624): 1394–1399.
ScienceNews (2020): A cat appears to have caught the coronavirus, but it’s complicated. https://www.sciencenews.org/article/
cats-animals-pets-coronavirus-covid19 (accessed 02.02.2021).
Shi J, Wen Z, Zhong G, Yang H, Wang C, Huang B, Liu R,
He X, Shuai L, Sun Z, Zhao Y, Liu P, Liang L, Cui P,
Wang J, Zhang X, Guan Y, Tan W, Wu G, Chen H, Bu Z
(2020): Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-coronavirus 2. Science 368(6494): 1016–1020.
Shim E, Tariq A, Choi W, Lee Y, Chowell G (2020): Transmission potential and severity of COVID-19 in South Korea. Int J Infect Dis 93: 339–344.
Sia SF, Yan LM, Chin AW, Fung K, Choy KT, Wong AY, Yen HL (2020): Pathogenesis and transmission of SARS-CoV-2 in golden hamsters. Nature 583(7818): 834–838.
Singh K, Hsu L-Y, Villacian JS, Habib A, Fisher D, Tambyah PA (2003): Severe acute respiratory syndrome: lessons from Singapore. Emerg Infect Dis 9(10): 1294–1298.
Sit THC, Brackman CJ, Ip SM, Tam KWS, Law PYT, To EMW, Yu VYT, Sims LD, Tsang DNC, Chu DKW, Perera RAPM, Poon LLM, Peiris M (2020): Infection of dogs with SARS-CoV-2. Nature 586(7831): 776–778.
Sridhar S, To KKW, Chan JFW, Lau SKP, Woo PCY, Yuen K-Y (2015): A systematic approach to novel virus discovery in
emerging infectious disease outbreaks. J Mol Diagn 17(3): 230–241.
Steinbrook R (2020): Contact Tracing, Testing, and Control of COVID-19-Learning From Taiwan. JAMA Intern Med 180(9): 1163–1164.
Su S, Wong G, Shi W, Liu J, Lai ACK, Zhou J, Liu W, Bi Y, Gao GF (2016): Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses. Trends Microbiol 24(6): 490–502.
Tsang KW, Ho PL, Ooi GC, Yee WK, Wang T, Chan-Yeung M, Lam WK, Seto WH, Yam LY, Cheung TM, Wong PC, Lam B, Ip MS, Chan J, Yuen KY, Lai KN (2003): A cluster of cases of severe acute respiratory syndrome in Hong Kong. N Engl J Med 348(20): 1977–1985.
Tu C, Crameri G, Kong X, Chen J, Sun Y, Yu M, Xiang H, Xia X, Liu S, Ren T, Yu Y, Eaton BT, Xuan H, Wang L-F (2004): Antibodies to SARS coronavirus in civets. Emerg Infect Dis 10(12): 2244–2248.
Varia M, Wilson S, Sarwal S, McGeer A, Gournis E, Galanis E, Henry B (2003): Investigation of a nosocomial outbreak of severe acute respiratory syndrome (SARS) in Toronto, Canada. CMAJ 169(4): 285–292.
Wang C, Zheng X, Gai W, Zhao Y, Wang H, Wang H, Feng N, Chi H, Qiu B, Li N, Wang T, Gao Y, Yang S, Xia X (2017): MERS-CoV virus-like particles produced in insect cells induce specific humoural and cellular imminity in rhesus macaques. Oncotarget 8(8): 12686–12694.
Wang Hongxin, Shao J, Luo X, Chuai Z, Xu S, Geng M, Gao Z (2020): Wildlife consumption ban is insufficient. Science 367(6485): 1435.
Wang Leyi, Mitchell PK, Calle PP, Bartlett SL, McAloose D, Killian ML, Yuan F, Fang Y, Goodman LB, Fredrickson R, Elvinger F, Terio K, Franzen K, Stuber T, Diel DG, Torchetti MK (2020): Complete Genome Sequence of SARS-CoV-2 in a Tiger from a U.S. Zoological Collection. Microbiol Resour Announc 9(22): e00468-20.
Wang M, Jing H, Xu H, Jiang X, Kan B, Liu Q, Wan K, Cui B, Zheng H, Cui Z, Yan M, Liang W, Wang H, Qi X, Li Z, Li M, Chen K, Zhang E, Zhang S, Hai R, Yu D, Xu J (2005): Surveillance on severe acute respiratory syndrome associated coronavirus in animals at a live animal market of Guangzhou in 2004. Zhonghua Liu Xing Bing Xue Za Shi 26(2): 84–87.
Weingartl HM, Copps J, Drebot MA, Marszal P, Smith G, Gren J, Andova M, Pasick J, Kitching P, Czub M (2004): Susceptibility of pigs and chickens to SARS coronavirus. Emerg Infect Dis 10(2): 179–184.
Weiss S, Leibowitz J (2011): Coronavirus Pathogenesis. Adv Virus Res 81: 85–164.
Wentworth DE, Gillim-Ross L, Espina N, Bernard KA (2004): Mice susceptible to SARS coronavirus. Emerg Infect Dis 10(7): 1293–1296.
World Health Organization (WHO) (2003): Summary of probable SARS cases with onset of illness from 1 November 2002 to 31 July 2003. https://www.who.int/csr/sars/country/table2004_04_21/en/ (accessed 23.07.2020).
World Health Organization (WHO) (2020): WHO Director-General’s opening remarks at the media briefing on COVID-19 – 11 March 2020. https://www.who.int/dg/speeches/detail/who-director-general-s-opening-r… (accessed 04.08.2020).
Wu D, Tu C, Xin C, Xuan H, Meng Q, Liu Y, Yu Y, Guan Y, Jiang Y, Yin X, Crameri G, Wang M, Li C, Liu S, Liao M, Feng L, Xiang H, Sun J, Chen J, Sun Y, Gu S, Liu N, Fu D, Eaton BT, Wang LF, Kong X (2005): Civets are equally susceptible to experimental infection by two different severe acute respiratory syndrome coronavirus isolates. J Virol 79(4): 2620–2625.
Xu R-H, He J-F, Evans MR, Peng G-W, Field HE, Yu D-W, Lee C-K, Luo H-M, Lin W-S, Lin P, Li L-H, Liang W-J, Lin J-Y, Schnur A (2004): Epidemiologic clues to SARS origin in China. Emerg Infect Dis 10(6): 1030–1037.
Yin D, Gao Q, Zhu H, Li J. (2020): Public perception of urban companion animals during the COVID-19 outbreak in China. Health Place 65: 102399.
Zhao H (2020): COVID-19 drives new threat to bats in China. Science 367(6485): 1436.
Zhao Z, Zhang F, Xu M, Huang K, Zhong W, Cai W, Yin Z, Huang S, Deng Z, Wei M, Xiong J, Hawkey PM (2003): Description and clinical treatment of an early outbreak of severe acute respiratory syndrome (SARS) in Guangzhou, PR China. J Med Microbiol 52(Pt 8): 715–720.
Zhong NS, Zheng BJ, Li YM, Poon, Xie ZH, Chan KH, Li PH, Tan SY, Chang Q, Xie JP, Liu XQ, Xu J, Li DX, Yuen KY, Peiris, Guan Y (2003): Epidemiology and cause of severe acute respiratory syndrome (SARS) in Guangdong, People’s Republic of China, in February, 2003. Lancet 362(9393): 1353–1358.
Zhou H, Chen X, Hu T, Li J, Song H, Liu Y, Wang P, Liu D, Yang J, Holmes EC, Hughes AC, Bi Y, Shi W (2020): A Novel Bat Coronavirus Closely Related to SARS-CoV-2 Contains Natural Insertions at the S1/S2 Cleavage Site of the Spike Protein. Curr Biol 30(11): 2196-2203.e3.



Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2020-29-Kaiser.pdf (0.2 MB) herunterladen möchten
{kind=link}
{kind=link}