

Interlaboratory proficiency test for detection of Paenibacillus larvae, the causative agent of American foulbrood
Berliner und Münchener Tierärztliche Wochenschrift 133
DOI: 10.2376/1439-0299-2020-20
© Schlütersche Verlagsgesellschaft mbH & Co. KG. 2020
Publiziert: 11/2020
Zusammenfassung
Im Herbst 2018 wurde vom Nationalen Referenzlabor für Amerikanische Faulbrut in Zusammenarbeit mit dem LAVES – Institut für Bienenkunde Celle ein bundesweiter Ringversuch mit inkludierter Laborvergleichsuntersuchung durchgeführt, an dem insgesamt 26 Laboratorien teilgenommen haben, darunter auch eines aus dem Ausland. Es wurden identische Probenpanel, bestehend aus sieben Honig-Proben im Umfang von je mind. 30 g, in codierter Form an alle Teilnehmer mit der Bitte versandt, diese in einem vorgegebenen Zeitraum mit den für den Ringversuch vorgegebenen und mit den in der Routine angewandten Methoden zu untersuchen. Die Ergebnisse werden im Beitrag mitgeteilt und kommentiert. Sie bestätigen den guten Stand der AFB-Diagnostik und die Qualität der Einrichtungen in Deutschland, aber ebenso die Notwendigkeit einer Harmonisierung und Standardisierung der zur Diagnostik eingesetzten Materialien, insbesondere der Nährmedien
Summary
In autumn 2018, the German National Reference Laboratory for American foulbrood carried out a nationwide proficiency test with an included laboratory comparison test in cooperation with the LAVES – Institute for Apiculture, in which a total of 26 laboratories took part, including one from abroad. Identical sample panels, consisting of seven honey samples with a volume of at least 30 g each, were sent in coded form to all participants with the request that they be examined within a specified period of time using the methods prescribed for the proficiency test and the methods used in the routine of the laboratory. The results are shown and commented in the article. They confirm the good status of AFB diagnostics and the quality of the facilities in Germany, but also the need for harmonization and standardization of the materials used for diagnostics, especially the culture media.
Einleitung
Die Amerikanische Faulbrut (AFB) ist weltweit die gravierendste Brutkrankheit der Honigbiene (Apis mellifera; Forsgren et al. 2018). Sie ist ein bedeutender Risikofaktor für Völkerverluste in Europa (Chauzat et al. 2013, Jacques et al. 2017) und in der gesamten Europäischen Union eine anzeigepflichtige Bienenseuche. In Deutschland waren in den letzten 20 Jahren durchschnittlich 246 Bienenstände pro Jahr von einem Ausbruch der AFB betroffen (Tiergesundheitsjahresbericht 2018). Wird diese Brutkrankheit in Deutschland an einem Bienenstand festgestellt, kommt es für die betroffene Imkerei sowie die umliegenden Bienenstände zu erheblichen Einschränkungen durch die seitens der Bienenseuchen-Verordnung (BienSeuchV) zu ergreifenden Maßnahmen. Der Erreger, Paenibacillus larvae, ist ein grampositives, peritrich begeißeltes, sporenbildendes Bakterium (Genersch et al. 2006). Nur die Sporen sind infektiös, bleiben dies aber über Jahrzehnte, da sie ausgesprochen widerstandfähig sind (Genersch 2010). Von P. larvae wurden verschiedene ERIC Genotypen beschrieben, die alle pathogen sind, sich aber in ihrer Virulenz unterscheiden (Genersch et al. 2006, Ashiralieva und Genersch 2006). Die beiden Genotypen ERIC I und ERIC II werden weltweit regelmäßig aus an AFB erkrankten Bienenvölkern isoliert (Schäfer et al. 2014, Morrissey et al. 2015). Auf Volksebene ist ERIC I virulenter als ERIC II, was dazu führt, dass Infektionen mit P. larvae des Genotyps ERIC II häufig erst spät erkannt werden (Rauch et al. 2009). Die Untersuchung auf Sporen des Erregers P. larvae im Futterkranz ist eine ausgezeichnete Methode zur Diagnose und Früherkennung der AFB und hat gezeigt, dass der Erreger nicht ubiquitär ist (von der Ohe und Dustmann 1997). Auf Grundlage der Festlegungen in § 27(3) des Gesetzes zur Vorbeugung vor und Bekämpfung von Tierseuchen (Tiergesundheitsgesetz - TierGesG) obliegt es dem Nationalen Referenzlabor (NRL) für Bienenkrankheiten Ringversuche mit den von den zuständigen Behörden der Bundesländer mit der Untersuchung anzeigepflichtiger Bienenseuchen beauftragten Laboratorien durchzuführen. Dementsprechend hat das NRL für Bienenkrankheiten in Zusammenarbeit mit dem LAVES – Institut für Bienenkunde Celle einen Ringversuch mit inkludierter Laborvergleichsuntersuchung (LVU) zum Nachweis von P. larvae-Sporen aus Futterproben durchgeführt. Während für den Ringversuch eine genaue Vorgehensweise vorgegeben war, inklusive der für die Untersuchung verwendeten Materialien, sollten die teilnehmenden Labore in der LVU die Methodik anwenden, welche jeweils routinemäßig angewendet wird. Wie in der amtlichen Methodensammlung des Friedrich-Loeffler-Instituts beschrieben, werden alle Proben vor dem Beimpfen der Nährmedien erhitzt, um vegetative Stadien von Bakterien und Pilzen weitgehend abzutöten. Da einige P. larvae-Stämme sensitiver auf eine Hitzebehandlung reagieren als andere und gezeigt wurde, dass insbesondere ERIC II-Stämme nach einer Hitzebehandlung im Wachstum reduziert waren (Forsgren et al. 2008), sollte eine P. larvae-Probe des Genotyps ERIC II (DSM 17237), im Ringversuch mit und ohne Hitzebehandlung untersucht werden. Außerdem haben die langjährigen Erfahrungen der P. larvae Diagnostik in Deutschland gezeigt, dass insbesondere die zur Anzucht verwendeten Nährmedien einen Einfluss auf die Germination der Sporen und das Wachstum des Erregers haben können. Um diesen Faktor zu prüfen, wurde parallel zum Ringversuch, in welchem die Verwendung eines bestimmten Nährmediums vorgeschrieben war, eine LVU durchgeführt. Im Artikel werden die Ergebnisse beider Untersuchungen dargestellt und diskutiert.
Material und Methoden
Insgesamt haben 25 Laboratorien (meist Untersuchungsämter) aus ganz Deutschland am Ringversuch mit inkludierter LVU teilgenommen, sowie ein Laboratorium eines Nachbarlandes.
Top Job:
Probenmaterial
Am 10.10.2018 wurden an alle Teilnehmer jeweils sieben Proben im Umfang von je mind. 30 g versandt, welche zwei Negativkontrollen und fünf mit P. larvae-Sporen belastete, positive Proben umfassten. Grundlage für die Herstellung der Proben waren zwei Honige (A & B) des LAVES – Institut für Bienenkunde in Celle aus dem Jahr 2018, die frei von P. larvae-Sporen waren. Negative Proben: „259“ (A) und „461“ (B). Honig B wurde aufgrund der niedrigeren Fremdkeimbelastung zur Herstellung aller positiven Proben herangezogen. Für die Herstellung der ERIC I Proben wurde ein Honig aus den 1990er-Jahren mit sehr hoher P. larvae-Sporenbelastung (C) ausgewählt: „166“ (10 g C + 1.750 g B) und „746“ (20 g C + 1.750 g B). Für die Herstellung der ERIC II Proben wurden zwei Fremdhonige (D & E) und der original RSK16-Sporensuspension-Honig (F) verwendet (P. larvae, DSM 17237): „348“ (450 g D + 1.750 g B), „599“ (E) und „658“ (1,33 g F + 1.750 g B). Alle Honigproben wurden in jeweils einem Ansatz homogenisiert und Aliquots von mind. 30 g abgewogen. Die Bestimmung des ERIC-Genotyps erfolgte in einer repPCR mittels ERIC-Primern (Genersch und Otten 2003). Alle Proben wurden bis zum Versand bei Umgebungstemperatur gelagert und erreichten auf dem Postweg nach maximal zwei Tagen alle Teilnehmer, welche nach Erhalt der Proben maximal einen Monat Zeit für die Bearbeitung der Proben hatten.
Untersuchungsverlauf
Für den Ringversuch wurden die folgenden Methoden vorgegeben. Die Proben werden maximal einen Monat bei Raumtemperatur gelagert. Um die Proben leicht verarbeiten zu können, werden diese über Nacht bei 37 °C ± 2 °C erwärmt und anschließend ohne weitere Erwärmung durch Rühren homogenisiert. Von jeder Probe werden 5,0 g ± 0,1 g in ein Reagenzglas eingewogen. Jede Probe wird 1:2 (v/v) mit sterilem entmineralisiertem Wasser (5,0 ml ± 0,1 ml) verdünnt und mithilfe eines Schüttlers bzw. Rührers homogenisiert. Anschließend werden alle sieben Proben gleichzeitig im auf 90 °C ± 2 °C vorgeheizten Wasserbad 6 min ± 20 s erhitzt (keine zusätzlichen Proben werden gleichzeitig erhitzt), um sie anschließend vor der weiteren Verarbeitung wieder auf Raumtemperatur abkühlen zu lassen. Vor dem Beimpfen der Nährbodenplatten werden die Proben mithilfe eines Schüttlers bzw. Rührers homogenisiert. Von jeder Probe werden je 200 µl auf jeweils drei Nährbodenplatten (alle Labore dürfen ausschließlich Columbia-Schafblut Agar (CSA) Nährmedienplatten „PB5008“ von Oxoid verwenden) pipettiert und mit einem sterilen Drigalskispatel ausplattiert. Zusätzlich wurde die Probe „658“ ein weiteres Mal wie oben beschrieben, jedoch ohne Erhitzung im Wasserbad untersucht. Die Inkubation der Nährbodenplatten erfolgte bei 37 °C ± 2 °C aerob für 6 Tage im Brutschrank. Die Auswertung der Anzahl koloniebildender-Einheiten (= KBE) des P. larvae erfolgt nach drei und nach sechs Tagen.
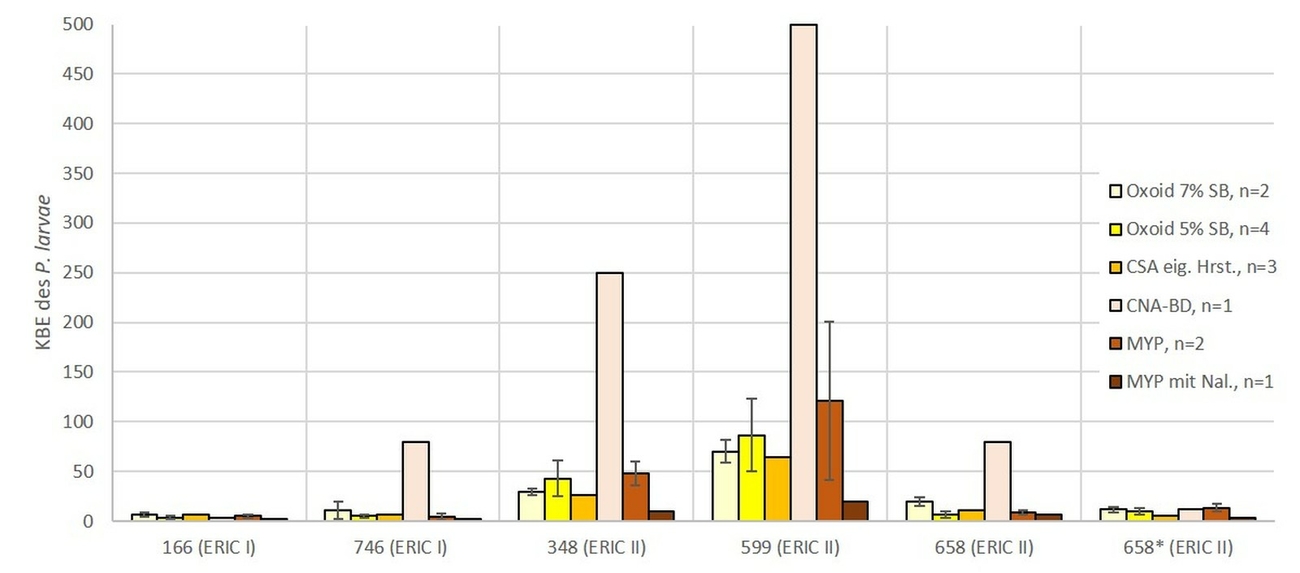
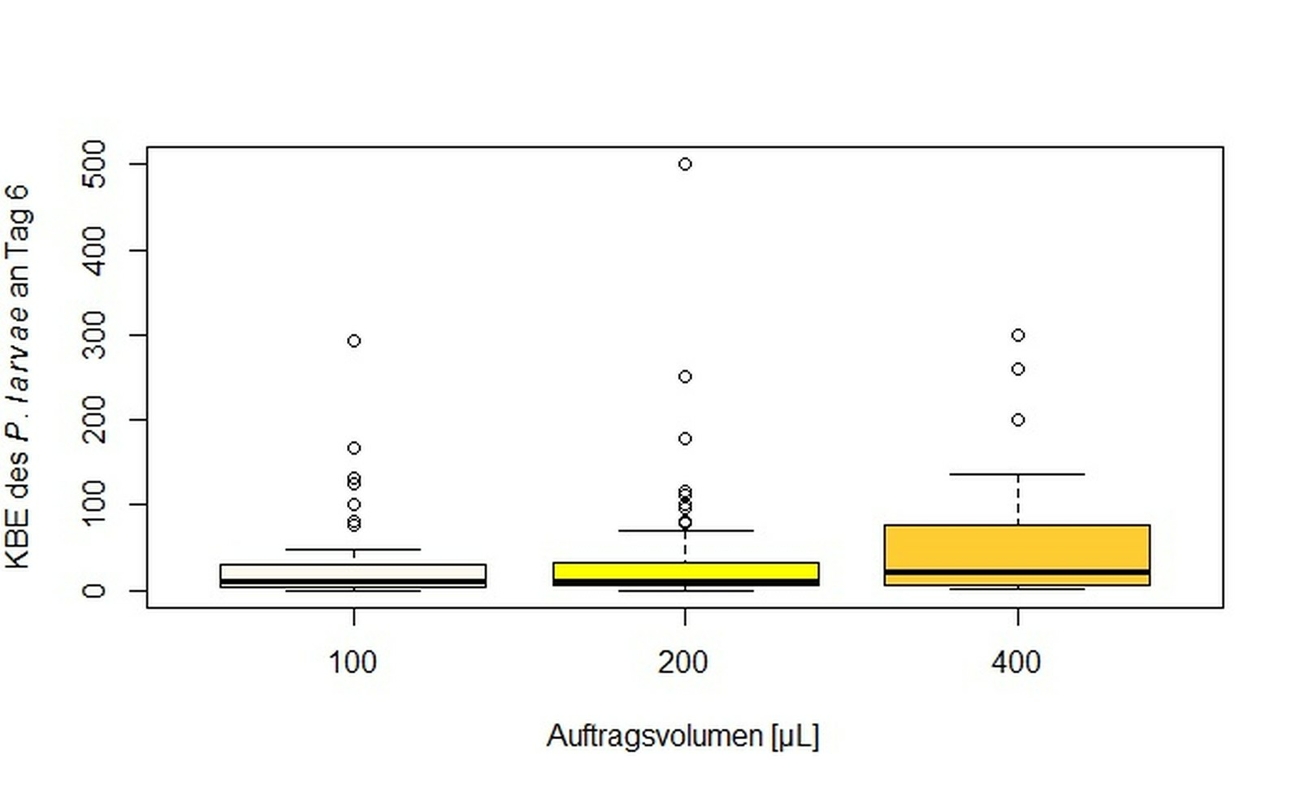
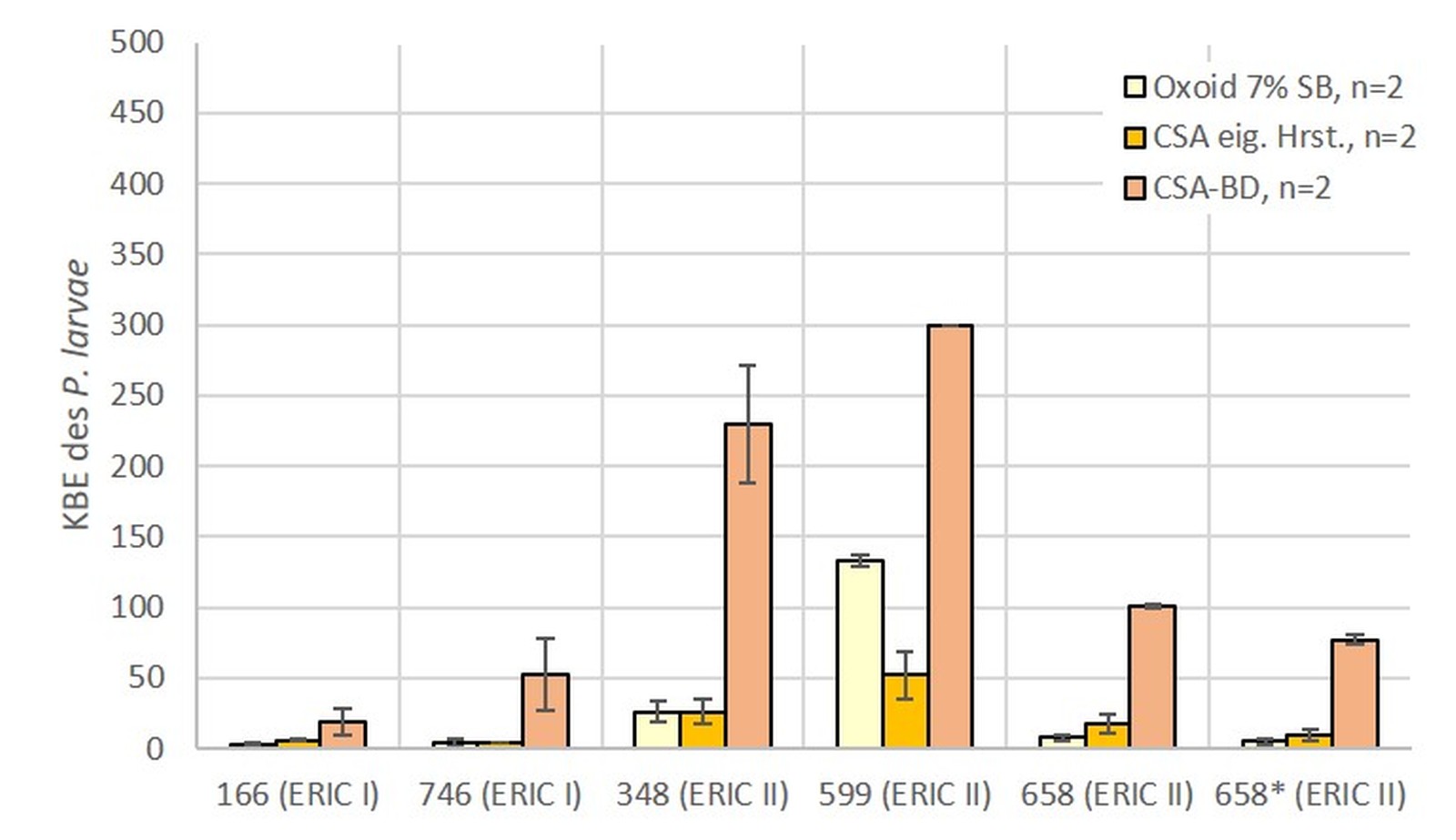
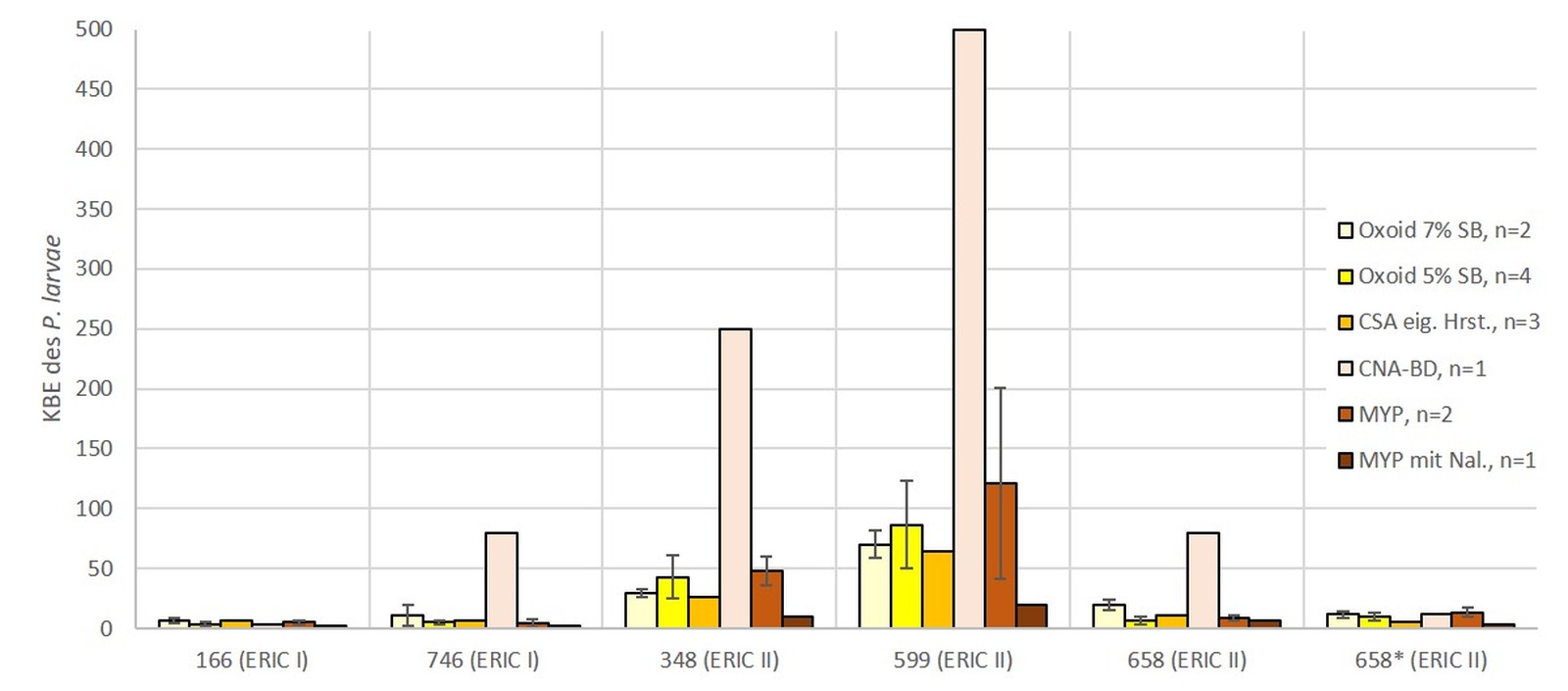
Bei der LVU wurde mit den in den Laboratorien jeweils routinemäßig anwendeten Methoden gearbeitet (Tab. 1). Es wurden unter anderem acht unterschiedliche Nährmedientypen (CSA von Oxoid mit 5 % (v/v) (n = 5) und mit 7 % (v/v) Schafsblut (n = 9), CSA aus eigener Herstellung (n = 5), CSA aus eigener Herstellung mit Zusatz von Nalidixinsäure (n = 3), CSA von Becton Dickinson (n = 2), CNA (mit Zusatz von Nalidixinsäure [10 mg/l] und Colistin [10 mg/l]) von Becton Dickinson (n = 1), sowie MYPGP aus eigener Herstellung, mit (n = 2) und ohne (n = 3) Zusatz von Nalidixinsäure verwendet. Auch wurden in der LVU drei unterschiedliche Volumina zur Inokulation der Nährböden genutzt: 100 µl, 200 µl und 400 µl (Tab. 1).
Auswertung
Für den Test auf Normalverteilung wurden die Proben mithilfe des Shapiro-Wilk-Tests untersucht, für Daten mit p-Werten > 0,05 konnte die Normalverteilung bestätigt werden, da sich keine signifikanten Unterschiede zeigten, eine andere Verteilung zu bevorzugen. Zusätzlich wurden die Ergebnisse, die im Ringversuch für Probe „658“ mit und ohne Hitzebehandlung ermittelt wurden, mithilfe des ungepaarten t-Tests (Welch-Test) untersucht. In der LVU wurde zum einen die Auswahl der eingesetzten Nährmedien und zum anderen die Menge der pro Platte aufgetragenen Suspension für einen Vergleich des Wachstums von KBE des P. larvae herangezogen. Die Daten wurden mittels generalisiertem linearem Model unter einer Gauß Verteilung analysiert und paarweise unter Berücksichtigung der least-square means verglichen. Alle Daten wurden mithilfe von R/R-Studio und dem R Package „lsmeans“ erhoben (Lenth 2016, R Core Team 2019, RStudio Team 2019).
Ergebnisse
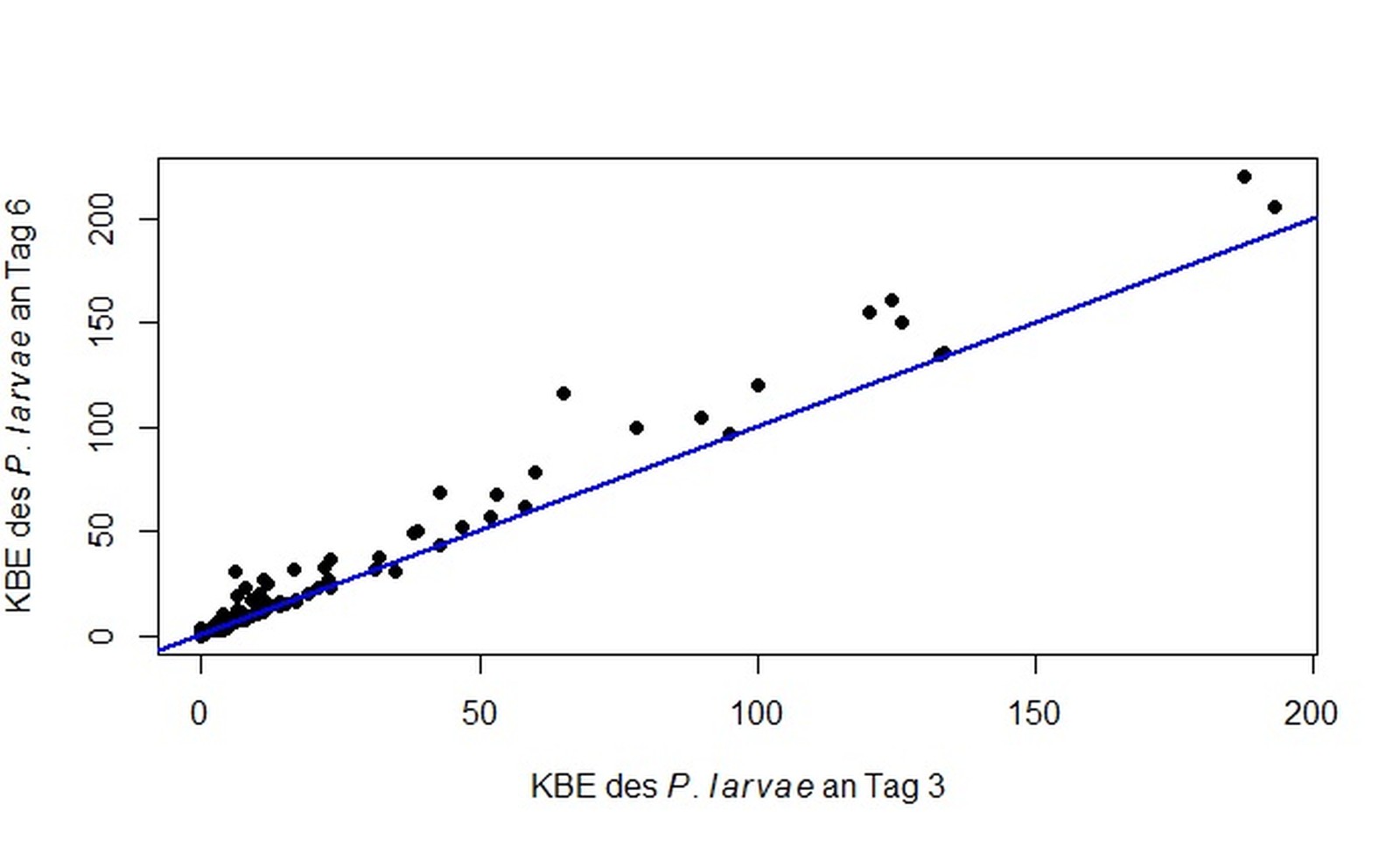
Im Ringversuch wurden von allen teilnehmenden Laboratorien nach sechs Tagen in der Regel gleich viele bzw. eine höhere Anzahl an KBE des P. larvae erfasst als nach drei Tagen (Abb. 1). Lediglich ein Labor zeigte für eine Probe nach drei Tagen eine höhere Anzahl KBE des P. larvae als nach sechs Tagen, was auf das stärkere Fremdkeimwachstum nach sechs Tagen zurückzuführen ist (Mitteilung des entsprechenden Labors). In drei der teilnehmenden Einrichtungen konnte für jeweils eine andere Probe nach drei Tagen noch kein Wachstum gezeigt werden. Während zwei dieser Einrichtungen für die jeweilige Probe nach sechs Tagen KBE des P. larvae nachweisen konnten, konnte ein Labor auch nach Tag sechs für die insgesamt am niedrigsten belastete Probe „166“ kein positives Ergebnis vorweisen. Bis auf Probe „746“ (p 0,001), waren die Ergebnisse normalverteilt: „166“ (p = 0,24), „348“ (p = 0,75), „599“ (p = 0,46), „658“ (p = 0,77) und „658*“ (p = 0,59). Im Median (± Standardabweichung) wurden für Probe „166“ nach 6 Tagen Inkubation 5,5 ± 3,2 KBE des P. larvae ermittelt. Probe „348“ zeigte einen Median von 32,0 ± 16,2 KBE des P. larvae, Probe „599“ einen Median von 100,0 ± 57,5 KBE des P. larvae, der Median der Probe „658“ lag bei 10,0 ± 5,3 KBE des P. larvae und der von Probe „746“ bei 7,0 ± 15,3 KBE des P. larvae. Die Probe „658“ kam ohne Hitzebehandlung auf einen Median von 8,5 ± 4,9 KBE des P. larvae und zeigte somit ein signifikant stärkeres Wachstum (p = 0,032; Abb. 2), wenn diese vor der Beimpfung der Nährbodenplatten im vorgeheizten Wasserbad bei 90 °C ± 2 °C für 6 min ± 20 s erhitzt wurde.
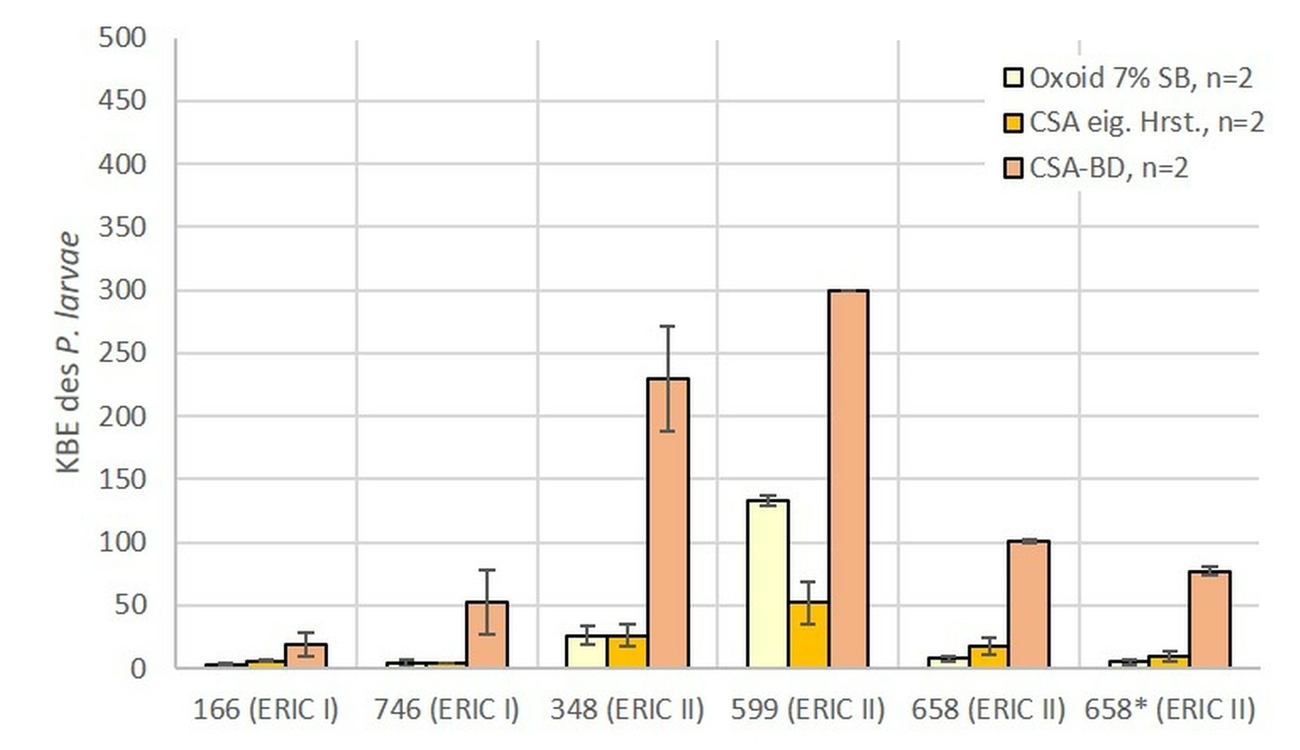
Von den fünf Nährmedien, welche in der LVU beim eingesetzten Volumen von 100 µl genutzt wurden (Abb. 3), zeigte der MYPGP Agar mit Nalidixinsäure signifikant stärkeres Wachstum gegenüber dem MYPGP Agar ohne Zusatz von Nalidixinsäure (p = 0,0175) und den beiden CSA Nährmedien von Oxoid (p = 0,003). Bei den sechs unterschiedlichen Nährmedien, welche unter Verwendung von 200 µl Suspension pro Nährmedienplatte in der LVU eingesetzt wurden, konnten auf den CNA Platten von Becton Dickinson signifikant mehr KBE des P. larvae nachgewiesen werden (p-Werte zwischen 0,0002 bis 0,046), als auf allen anderen Nährmedienplatten (Abb. 4). Unter den drei verschiedenen Nährmedienplatten, bei welchen in der LVU 400 µl Sporensuspension verwendet wurden, war die Anzahl der KBE des P. larvae auf dem CSA Agar von Becton Dickinson signifikant höher als bei den beiden anderen Nährmedien (p = 0,0002 für CSA und p = 0,001 für Oxoid) (Abb. 5). Die beiden Nährmedien von Oxoid wurden für die gesamte Auswertung zusammengefasst, da sich in den Ergebnissen keine signifikanten Unterschiede zeigten (p = 1). Der MYPGP-Agar mit Zusatz von Nalidixinsäure zeigte in einem der teilnehmenden Labore eine sehr gute Sensitivität (Abb. 3). Im Vergleich dazu, war die Sensitivität mit dem gleichen Agar-Typ in einem anderen Labor sehr niedrig (Abb. 4), obwohl hier das doppelte Volumen pro Platte verwendet wurde.
Vergleicht man die Anzahl der in der LVU im Durchschnitt gewachsenen KBE des P. larvae für die jeweiligen Volumina, die zur Inokulation verwendet wurden, kann kein signifikanter Unterschied zwischen den Volumina gezeigt werden (Abb. 6). Fasst man diese Daten zusammen und vergleicht alle verwendeten Nährmedien miteinander ist festzustellen, dass CSA und CNA von Becton Dickinson gegenüber allen anderen Medien eine signifikant höhere Sensitivität für P. larvae zeigten (Tab. 2).
Diskussion
Insgesamt ist festzustellen, dass alle Teilnehmer P. larvae erfolgreich nachweisen konnten. Zum Teil zeigten sich zwischen den Laboratorien große Unterschiede in den für die einzelnen Proben jeweils ermittelten KBE des P. larvae. Dies trifft sowohl auf die LVU zu, als auch auf den Ringversuch, in welchem die Vorgehensweise, inklusive eines Mediums vom gleichen Hersteller, vorgegeben war. Dies ist vor allem auf das Fremdkeimwachstum zurückzuführen, welches die Auswertung in vielen Fällen im Ringversuch und in der LVU problematisch gestaltete (entsprechende Anmerkungen wurden bei Übermittlung der Ergebnisse von acht der teilnehmenden Laboratorien mitgeteilt). Obwohl Aliquots derselben, homogenisierten Ansätze verwendet wurden, wird die Zahl der ermittelten KBE des P. larvae auch generell durch die Variabilität in der ohnehin niedrigen Keimungsrate der Sporen unterschiedlicher P. larvae-Stämme beeinflusst (Forsgren et al. 2008). So ermittelten Forsgren et al. (2008) durch den Vergleich der mittels Zählkammer ermittelten Sporenmenge in einer Suspension und der daraus gekeimten Anzahl von KBE bei fünf untersuchten P. larvae-Stämmen der Genotypen ERIC I und II eine Keimungsrate von nur ca. 10 %.
Auch die bei Probe „166“ insgesamt niedrige Anzahl der ermittelten KBE des P. larvae, erklären das Auftreten eines negativen Ergebnisses im Ringversuch für eben diese, eigentlich positive Probe. Aufgrund der statistischen Möglichkeit des Auftretens eines negativen Resultats, wurde dieses negative Ergebnis in der Bewertung des Ringversuchs nicht als falsch-negatives Kriterium angesehen.
Von den methodischen Abweichungen in der LVU (Tab. 1) eigneten sich zum einen die Auswahl der eingesetzten Nährmedien und zum anderen die Menge der pro Platte aufgetragenen Suspension für einen Vergleich.
Im Allgemeinen zeigen die CSA und CNA-Agare von Becton Dickinson das beste Wachstum des Zielorganismus (Tab. 2; Abb. 4 und 5). Auch der MYPGP-Agar mit Zusatz von Nalidixinsäure zeigte ein sehr gutes Wachstum in einem Labor (Abb. 3). Allerdings waren im Vergleich dazu, die Ergebnisse mit dem gleichen Agar-Typ in einem anderen Labor sehr niedrig (Abb. 4), obwohl hier das doppelte Volumen pro Platte verwendet wurde. Ein Grund hierfür könnten Unterschiede in der Herstellung der Nährbodenplatten sein, welche zu dem komplexen Nährmedien zählen und die jeweils in hauseigenen Nährbodenküchen erfolgte.
Für den Vergleich der Ergebnisse hinsichtlich des pro Platte aufgetragenen Suspensionsvolumens zeigte sich kein signifikanter Unterschied, aber die rechnerische Steigerung in der Anzahl der zu erwartenden KBE des P. larvae bei höherem Volumen bildet sich tendenziell ab (Abb. 6), insbesondere die Probe „599“ zeigt eine stetige Zunahme der KBE des P. larvae mit höherem Volumen. Hinsichtlich der Anwendung im Laborbetrieb ist jedoch das Volumen von 200 µl dem von 400 µl vorzuziehen, da das kleinere Volumen besser von den Nährmedien aufgenommen wird.
Zusätzlich muss hier noch angemerkt werden, dass in der LVU neben den verwendeten Nährmedien und der Menge der pro Platte aufgetragenen Suspension, auch weitere Parameter in der Methodik zwischen den teilnehmenden Laboratorien teilweise variierte (siehe Tab. 1).
Da gezeigt wurde, dass einige P. larvae-Stämme sensitiver auf eine Hitzebehandlung reagieren als andere und insbesondere ERIC-II-Stämme nach einer Hitzebehandlung im Wachstum reduziert waren (Forsgren et al. 2008), untersuchten wir Probe „658“ (ERIC II) im Ringversuch mit und ohne Hitzebehandlung. Wir konnten das Ergebnis von Forsgren et al. (2008), dass insbesondere ERIC-II-Stämme nach einer Hitzebehandlung im Wachstum reduziert waren, nicht bestätigen. Diese Probe zeigte mit Hitzebehandlung ein signifikant höheres Wachstum als ohne Hitzebehandlung (Abb. 2). Dies ist vor allem darauf zurückzuführen, dass Forsgren et al. (2008) die Hitzebehandlung länger (10 min) und mit wesentlich geringerem Volumen (100 µl im Vergleich zu 10 ml) durchführten (Forsgren E., persönliche Mitteilung). Somit sehen wir keine Notwendigkeit die amtliche Methodensammlung hinsichtlich der Erhitzung der Proben vor dem Beimpfen der Nährmedien zu verändern, da die Erhitzung von 10 ml Suspension bei 90 °C für 6 min keinen negativen Einfluss auf die Keimungsrate der untersuchten Probe des Genotyps ERIC II zeigte.
Fazit
In der Regel war das Wachstum von P. larvae auf Nährböden von Becton Dickinson um ein Vielfaches besser als auf den anderen verwendeten Nährböden. Das Volumen von 200 µl Suspension pro Nährbodenplatte reicht für den semi-quantitativen Nachweis des AFB-Erregers aus und zeigt in der Tendenz ein erhöhtes Wachstum von KBE des P. larvae im Vergleich zu 100 µl pro Platte. In den teilnehmenden Untersuchungseinrichtungen sollten die Untersuchungsmethoden entsprechend harmonisiert und standardisiert werden, um die in der Routinediagnostik erzielten Ergebnisse in Zukunft besser zwischen den Untersuchungseinrichtungen vergleichen zu können.
Acknowledgement
Wir danken allen teilnehmenden Laboratorien.
Ethische Anerkennung
Der Artikel beinhaltet keine Versuchstier-Studien, welche von den Autoren durchgeführt wurden.
Conflict of interest
Es bestehen keine geschützten, finanziellen, beruflichen oder anderen persönlichen Interessen an einem Produkt, Service und/oder einer Firma, welche die im oben genannten Manuskript dargestellten Inhalte oder Meinungen beeinflussen könnten.
Funding
Diese Arbeit wurde unterstützt vom Friedrich-Loeffler-Institut, Bundesforschungsinstitut für Tiergesundheit und dem LAVES – Institut für Bienenkunde Celle. Die Autoren versichern, dass sie Daten hierzu auf begründete Nachfrage hin bereitstellt.
Korrespondenzadresse
Dr. Marc Schäfer
Nationales Referenzlabor für Bienenkrankheiten
Friedrich-Loeffler-Institut
Südufer 10
17493 Greifswald – Insel Riems
marc.schaefer@fli.de
Literatur
Ashiralieva A, Genersch E (2006): Reclassification, genotypes and virulence of Paenibacillus larvae, the etiological agent of American foulbrood in honeybees – a review. Apidologie 37: 411–420.
Chauzat M-P, Cauquil L, Roy L, Franco S, Hendrikx P, Ribière-Chabert M (2013): Demographics of the European Apicultural Industry. PLoS ONE 8(11): e79018.
Forsgren E, Stevanovic J, Fries I (2008): Variability in germination and in temperature and storage resistance among Paenibacillus larvae genotypes. Vet Microbiol 129: 342–349.
Forsgren E, Locke B, Sircoulomb F, Schäfer MO (2018): Bacterial diseases in honeybees. Curr Clin Microbiol Rep 5: 18–25.
Friedrich-Loeffler-Institut, Bundesforschungsinstitut für Tiergesundheit (2019): Tiergesundheitsjahresbericht 2018.
Genersch E (2010): American Foulbrood in honeybees and its causative agent, Paenibacillus larvae. J Inv Path 103: S10–S19.
Genersch E, Otten C (2003): The use of repetitive element PCR fingerprinting (rep-PCR) for genetic subtyping of German field isolates of Paenibacillus larvae subsp. larvae. Apidologie 34: 195–206.
Genersch E, Forsgren E, Pentikäinen J, Ashiralieva A, Rauch S, Kilwinski J, Fries I (2006): Reclassification of Paenibacillus larvae subsp. pulvifaciens and Paenibacillus larvae subsp. larvae as Paenibacillus larvae without subspecies differentiation. Int. J. Syst Evol Microbiol 56: 501–511.
Jacques A, Laurent M, EPILOBEE Consortium, Ribière-Chabert M, Saussac M, Bougeard S, Budge GE, Hendrikx P, Chauzat M-P (2017): A pan-European epidemiological study reveals honey bee colony survival depends on beekeeper education and disease control. PLoS ONE 12(3): e0172591.
Lenth RV (2016): Least-Squares Means: The R Package lsmeans. J Stat Softw 69(1): 1–33.
Morrissey BJ, Helgason T, Poppinga L, Fünfhaus A, Genersch E, Budge GE (2015): Biogeography of Paenibacillus larvae, the causative agent of American foulbrood, using a new multilocus sequence typing scheme. Environ Microbiol 17: 1414–24.
R Core Team (2019): R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/.
Rauch S, Ashiralieva A, Hedtke K, Genersch E (2009): Negative correlation between individual-insect-level virulence and colony-level virulence of Paenibacillus larvae, the etiological agent of American foulbrood of honeybees. Appl Environ Microbiol 75: 3344–3347.
RStudio Team (2019): RStudio: Integrated Development for R. RStudio, Inc., Boston, MA URL http://www.rstudio.com/.
Schäfer MO, Genersch E, Fünfhaus A, Poppinga L, Formella N, Bettin B, Karger A (2014): Rapid identification of differentially virulent genotypes of Paenibacillus larvae, the causative organism of American foulbrood of honey bees, by whole cell MALDI-TOF mass spectrometry. Vet Microbiol 170: 291–297.
Von der Ohe W, Dustmann JH (1997): Efficient prophylactic measures against American foulbrood by bacteriological analysis of honey for spore contamination. Am Bee J 137: 603–606.




Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2020-20-Schaefer.pdf (0.82 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2020-20-Schaefer-Tabelle1.pdf (0.17 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2020-20-Schaefer-Tabelle2.pdf (0.11 MB) herunterladen möchten

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}