

Untersuchung zur Ausscheidung ausgewählter pathogener, zoonotischer oder antimikrobiell resistenter Bakterien bei südamerikanischen Kameliden in Mitteldeutschland
Berliner und Münchener Tierärztliche Wochenschrift 135, 1–16
DOI: 10.2376/1439-0299-2021-21
© Schlütersche Fachmedien GmbH. 2022
Eingereicht: 26. August 2021
Akzeptiert: 13. Januar 2022
Publiziert: 02/2022
Summary
South American camelids (SAC) are increasingly popular in Germany. Although frequently in close contact to livestock and humans, data on the prevalence of epizootic and zoonotic bacteria and of bacteria exhibiting antimicrobial resistance (AMR) in SAC is sparse. Therefore, 94 composite faecal samples collected in 43 private SAC holdings in the German states Saxony, Thuringia and Saxony-Anhalt were tested for the presence of Clostridium difficile, Mycobacterium avium subsp. paratuberculosis (MAP), Salmonella spp. and Chlamydia spp. Escherichia coli served as an AMR-indicator and for detecting Shiga toxin (Stx)-producers (STEC). Chlamydial presence was also probed with 136 vaginal swabs and twelve placental tissue samples. Most owners (n = 41/43) replied to a questionnaire providing information on animals and husbandry conditions. The questionnaire covered 974 animals, among them 20 llamas, mostly kept in smallholdings with less than 20 animals (75%). Frequently (n = 30/41), SAC were co-housed with other animal species. Most holdings (76%, n = 31/41) welcomed visitors regularly. Clostridium difficile was detected in 8.5% (n = 8/94) of the faecal samples, Chlamydia spp. in 7.4% (n = 7/94) of the faecal samples, 4.4% (n = 6/136) of the vaginal swabs and 8.3% (n = 1/12) of the placental samples. MAP and Salmonella spp. were not detected. Samples from 28% of the holdings were stx-positive. STEC strains isolated belonged to eae-negative, non-O157 serovars. All E. coli isolates were resistant to at least five antibiotics with 46% possible ESBL-producers. Even though shedding of bacteria of public health concern by SAC at livestock-livestock and livestock-human interfaces was found to be less common, positive results indicate the need for establishing continuous surveillance measures.
Zusammenfassung
Neuweltkameliden (NWK) werden in Deutschland immer beliebter. Obwohl sie häufig in engem Kontakt zu Nutztieren und Menschen gehalten werden, ist wenig über die Prävalenz von bakteriellen Tierseuchen- und Zoonoseerregern oder von Bakterien mit antimikrobiellen Resistenzen (AMR) bei diesen Tieren bekannt. Daher wurden 94 Sammelkotproben von NWK aus 43 Privatbetrieben in Sachsen, Thüringen und Sachsen-Anhalt gesammelt und auf Clostridium difficile, Mycobacterium avium subsp. paratuberculosis (MAP), Salmonella spp. und Chlamydia spp. untersucht. Escherichia coli diente als Indikator für AMR und zum Nachweis von Shigatoxin (Stx)-Produzenten (STEC). Nach Chlamydien wurde auch in 136 Vaginalabstrichen und zwölf Plazentagewebeproben gesucht. Die meisten Besitzer (n = 41/43) beantworteten einen Fragebogen zu den Tieren und deren Haltungsbedingungen. Über den Fragebogen wurden 974 Tiere erfasst, darunter 20 Lamas, die meist in kleinen Betrieben mit weniger als 20 Tieren (75 %) gehalten wurden. Häufig (n = 30/41) wurden SAC zusammen mit anderen Tierarten gehalten. Die meisten Betriebe (76 %, n = 31/41) empfangen regelmäßig Besucher. Clostridium difficile wurde in 8,5 % (n = 8/94) der Kotproben, Chlamydia spp. in 7,4 % (n = 7/94) der Kotproben, 4,4 % (n = 6/136) der Vaginalabstriche und 8,3 % (n = 1/12) der Plazenta-Proben nachgewiesen. MAP und Salmonella spp. wurden nicht nachgewiesen. Siebzehn Kotproben aus 28 % der Betriebe waren stx-positiv. Aus diesen isolierte STEC-Stämme waren eae-negativ und gehörten zu Nicht-O157-Serovaren. Auf Antibiotikaresistenz untersuchte E. coli-Isolate waren gegen mindestens fünf Antibiotikaklassen resistent. Von diesen waren 46 % potenzielle ESBL-Produzenten. Auch wenn die Ergebnisse eine fehlende oder mäßige Verbreitung von bakteriellen Tierseuchen- und Zoonoseerregern bei NWK in Deutschland implizieren, sprechen die positiven Nachweise für die Notwendigkeit, nachhaltige Überwachungsmaßnahmen zu etablieren.
Introduction
South American camelids (SAC) are classified as four different species: alpacas (Vicugna pacos), vicuñas (Vicugna vicugna), llamas (Lama glama) and guanacos (Lama guanicoe) (Zarrin et al. 2020). In recent years, these animals have been introduced more and more to North America and Europe (Zarrin et al. 2020), including Germany (Neubert et al. 2021). SAC are held in full- or part-time commercial farming for their wool or for breeding (Halsby et al. 2017), but also for landscape management, for use in animal assisted therapy, for exhibition or as pets (Neubert et al. 2021). Their number in Germany was estimated to be roughly 15,000 animals in 2018 (Gauly 2019). However, it is certainly higher today, considering that the SAC count listed on the website of a German breeding organization, the Alpaka Zucht Verband Deutschland, increased by 16% from 12,458 in August 2020 (Neubert et al. 2021) to 14,458 in December 2021 (azvd.de; 10.12.2021).
SAC are recognized as carriers of different epizootic agents, including those causing notifiable diseases according to Commission Implementing Regulation (EU) 2018/1882, such as infection with epizootic hemorrhagic disease virus, with members of the Mycobacterium tuberculosis complex or with paratuberculosis, as well as zoonotic agents with the potential to harm humans. However, the current animal health status of SAC in Germany has not yet been systematically recorded (Neubert et al. 2021). Experience has shown that there are knowledge gaps among both animal owners and veterinarians, because there is still uncertainty concerning disease diagnosis and treatment (D’Alterio et al. 2006).
Camelids (suborder Tylopoda) differ taxonomically, physiologically and behaviorally from ruminants (suborder Ruminantia) (Fowler 2009). Moreover, camelids and ruminants diverge in their predisposition to infectious and parasitic diseases. For example, camelids are quite resistant to many ruminant diseases, such as Foot-and-mouth disease and bovine tuberculosis (TB) caused by Mycobacterium bovis (Halsby et al. 2017). However, infections with Mycobacterium avium subsp. paratuberculosis (MAP) or with Campylobacter fetus subsp. fetus, Chlamydia spp., Salmonella spp. and Brucella abortus can lead to disease, for example by causing abortions (Gidlewski et al. 2000, Tibary et al. 2006, Staples 2016, Halsby et al. 2017). Other important bacterial agents reportedly infecting SAC are Clostridium perfringens (types A and C enterotoxaemia), Clostridium septicum, Streptococcus equi subsp. zooepidemicus (Whitehead and Bedenice 2009, Staples 2016), and Mycoplasma haemolamae (Viesselmann et al. 2019). So far, alpacas kept in Germany have been investigated for the presence of zoonotic viruses, such us Cowpox (Prkno et al. 2017), Schmallenberg orthobunyavirus (Schulz et al. 2015) and Borna disease virus 1 (BoDV-1) (Schulze et al. 2020). BoDV-1 produces a highly variable spectrum of symptoms in NWK (Schulze et al. 2020) and can also serve as a sentinel species (Malbon et al. 2021).
Shiga toxin (Stx)-producing Escherichia coli (STEC), harboring stx1 and/or stx2 (Blankenship et al. 2021), are zoonotic bacteria that can cause mild to bloody diarrhea, hemorrhagic colitis, or even hemolytic uremic syndrome in humans (Gyles 2007). Ruminants are the major STEC reservoir, especially cattle (Gyles 2007). However, STEC was also detected in non-ruminant animals (Persad and LeJeune 2014) including alpacas and llamas (Mercado et al. 2004, Silvera et al. 2012, Halsby et al. 2017, Maturrano et al. 2018). In Peru, alpaca crias suffering from diarrhea infrequently carried stx2-encoding STEC with a low level of verocytotoxic activity, while the large majority (94.7%) shed enteropathogenic (EPEC) strains (Luna et al. 2012). Other studies did not report the presence of enterotoxigenic E. coli (ETEC) and EPEC (Whitehead and Bedenice 2009, Staples 2016). In SAC, E. coli has usually been reported as causing diarrhea, metritis, mastitis or as a pathogen commonly associated with dental abscessation (Whitehead and Bedenice 2009, Staples 2016, Gestrich et al. 2018). In neonatal and older crias, meningitis, meningoencephalitis and septicemia also occur (Staples 2016). Septicemia was the second most frequently diagnosed non-parasitic problem in the United Kingdom and E. coli caused 43.3% of these cases (Twomey et al. 2014). Commensal and pathogenic E. coli in faeces and sewage serve as indicator organisms for AMR in Gram-negative bacteria in human and veterinary public health as they can contain genetic elements with resistance determinants transferable to other enteric bacteria (EFSA and ECDC 2020). In one study from Peru, alpaca crias, with or without diarrhea, carried E. coli susceptible to most antibiotics, with the exception of nitrofurantoin, to which only few strains (8.3%) had retained susceptibility (Barrios-Arpi et al. 2016). However, other studies found 80% of the E. coli strains resistant to neomycin and 25% to oxytetracycline (Luna et al. 2012) or detected gentamicin resistance in 17 of 98 bacterial isolates (Gestrich et al. 2018).
SAC have been included as livestock species in the profoundly revised European Animal Health Law. Even though they represent potential sources of transmission of infectious agents to other livestock species and humans, because they are frequently kept on the same premises and humans are often in close contact with the animals, knowledge on the prevalence of infectious agents in SAC in Germany is sparse. We surveyed SAC in Central Germany for selected pathogenic and zoonotic bacteria, as well as for antimicrobial resistance in indicator E. coli, but also for population structure and husbandry conditions, to foster future zoonosis monitoring and preventive medicine measures.
Top Job:
Material and Methods
Sampling and questionnaire
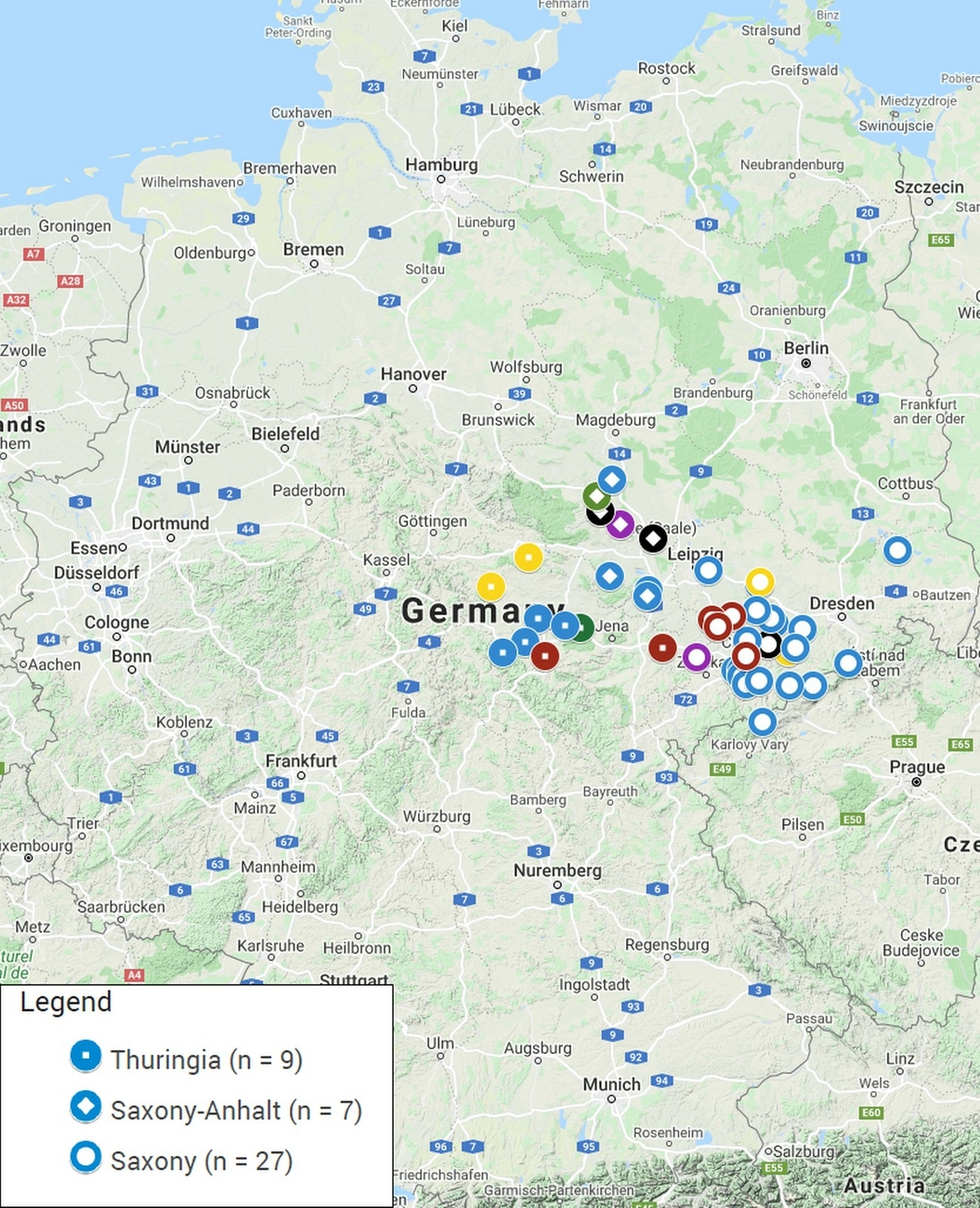
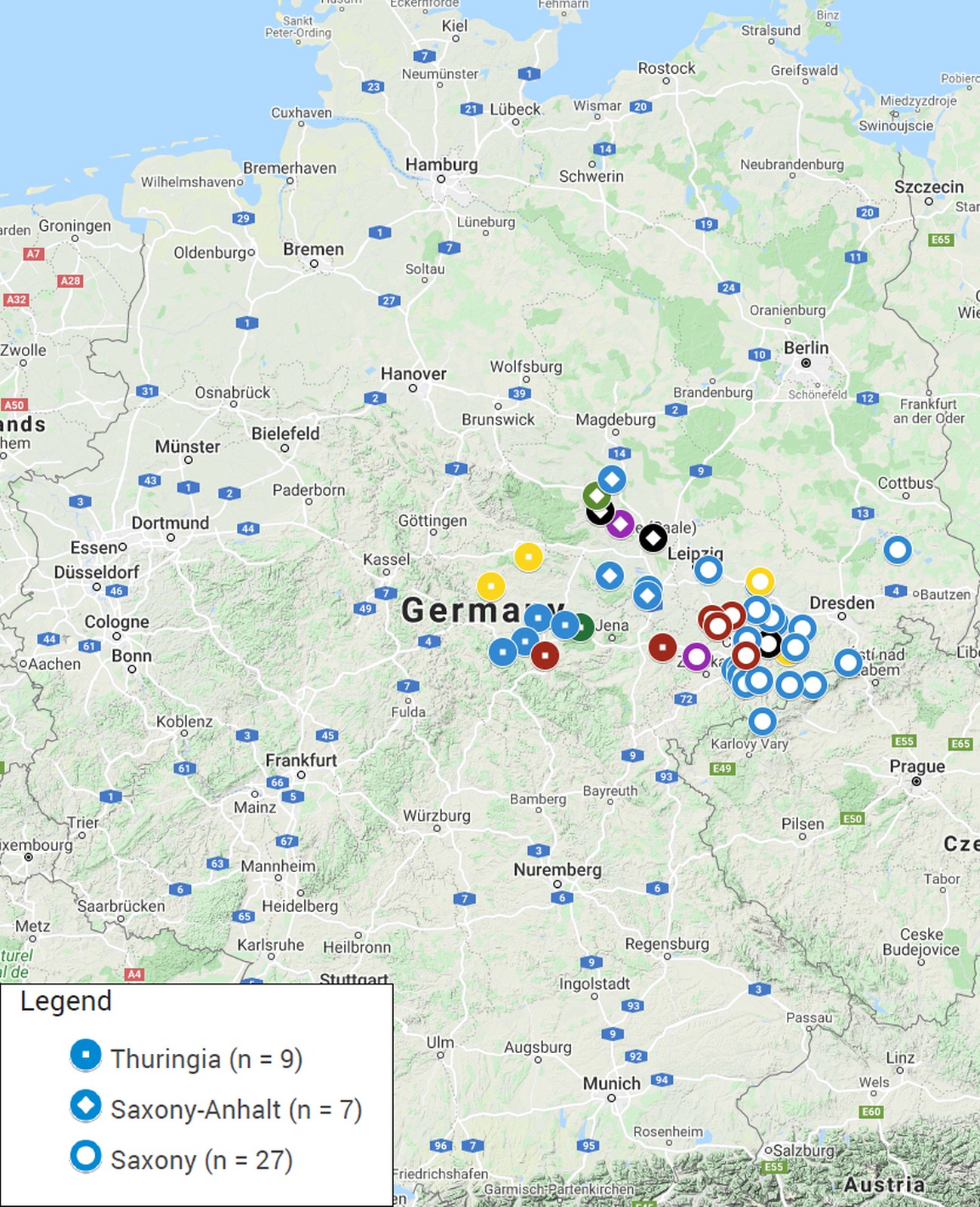
94 composite faecal samples from fresh droppings from alpaca and llama animal groups were collected on 43 farms or zoos located in the three German federal states (Saxony, Thuringia and Saxony-Anhalt) comprising the region Central Germany (Fig. 1, Supplementary Material 1). Faecal material was collected from pastures or pen floors in 100 ml containers and transported to the laboratory on ice. Vaginal swabs (FLOQSwabs, Copan, Brescia, Italy) were collected from 136 mares by a veterinarian, the tips cut off and placed in tubes containing 300 µl DNA stabilizing buffer (6 M Guanidin-HCl, 10 mM urea, 10 mM Tris-HCl, 20% v/v Triton X-100; pH 4.4). Placenta samples from twelve normal deliveries were processed on the day of arrival in the lab or stored at 5 °C for processing on the next day.
The study was carried out in strict accordance with German law for the care and use of experimental animals. Sampling of vaginal swabs was approved by the Thuringian State Office for Food Safety and Consumer Protection, Bad Langensalza, Germany (permit no. 22-2684-04-BFI-19-101), by the State Administrative Office of Saxony-Anhalt, Halle, Germany (permit no. 203.m-42502-3-865 FLI_G_Ü) and by the Saxony State Directorate, Chemnitz, Germany (permit no. 25-5131/475/8), respectively, depending on the locations of the holdings.
The owners received a cover letter explaining the purpose of the investigation and signed a declaration of consent according to Directive 95/46/EC (General Data Protection Regulation) for taking samples and analyzing them for scientific reasons and to ensure confidentiality. Owners also received a multiple-choice questionnaire with 50 questions divided in five sections: general information (farm and husbandry), animal transfer, breeding, health status and movement of people (Supplementary Material 2).
Screening for Clostridium difficile
Faecal samples were processed for the isolation of Clostridium difficile (C. difficile) as described previously (Schneeberg et al. 2012) using C. difficile moxalactam/norfloxacin agar plates (CDMN; Oxoid, Thermo Fisher Scientific, Waltham, MA, USA) and ChromID C. difficile agar (bioMérieux, Nürtingen, Germany) for direct plating (2–3 days) and TCDMN broth (CDMN broth supplemented with 0.1% sodium taurocholate) for enrichment (7–10 days). The culturing method after enrichment involved an ethanol shock (1 mL culture : 1 mL absolute ethanol; 30 min) for spore selection as described elsewhere (Schneeberg et al. 2012) followed by plating on CDMN and ChromID agar. Potential C. difficile colonies were selected based on their branched morphology and growth on CDMN (Oxoid) and ChromID C. difficile agar (bioMérieux). Strain isolation included three successive subcultures of single colonies. Bacterial DNA was prepared using the DNeasy Blood and Tissue kit (Qiagen, Hilden, Germany). Isolates were confirmed as C. difficile using cdd3-PCR as described (Zidaric et al. 2008).
Screening for Chlamydia
Homogenized pooled faecal samples, placenta tissue (50 mg each) and vaginal swabs were extracted using the High Pure PCR Template Preparation Kit (Roche Life Science, Penzberg, Germany) according to the manufacturer’s protocol with an elution volume of 200 µL. All samples were examined using a 23S-rRNA gene-based Chlamydiaceae-specific real-time PCR with a limit of detection of 5 copies/reaction (Ehricht et al. 2006). QuantiTect Multiplex PCR Master Mix (Qiagen) was used in a total volume of 15 µL with 300 nM of each primer, 200 nM probe and 2 µL template DNA. To test for PCR inhibition in DNA preparations, an internal amplification control was integrated in duplex PCR runs. Primers EGFP-1F (5’-GACCACTACCAGCAGAACAC-3’) and EGFP-10R (5’-CTTGTACAGCTCGTCCATGC-3’) at final concentrations of 400 nM each and probe EGFP-HEX (HEX-AGCACCCAGTCCGCCCTGAGCA-BHQ1) at a final concentration of 200 nM were used together with 500 copies of a plasmid template per reaction (Intype IC-DNA, Indical Bioscience, Leipzig, Germany) to generate and detect a 177 bp amplicon (Hoffmann et al. 2006). The samples were tested in duplicate on a Bio-Rad CFX96 PCR instrument (Bio-Rad, Feldkirchen, Germany) with the following cycling conditions: 95 °C for 10 min and 45 cycles with 95 °C for 15 s and 60 °C for 1 min. A mean threshold cycle (Ct) value of 38 was considered positive. A 100 ifu/µl equivalent of Chlamydia psittaci (C. psittaci) strain DC15 DNA was used as positive control and water instead of template DNA as negative control. 23S-PCR-positive samples were further characterized by species-specific (Chlamydia abortus [C. abortus], Chlamydia pecorum [C. pecorum], C. psittaci) real-time PCR assays (Pantchev et al. 2010) with primer concentrations of 800 nM each and probe concentrations of 200 nM. Mastermix and PCR cycling conditions were as described above.
Screening for Mycobacterium avium subsp. paratuberculosis (MAP)
Pooled faecal samples were thoroughly homogenised by manual stirring. Three grams of faeces were transferred to a plastic bag containing 30 mL 0.75% hexadecyl pyridinium chloride. The samples were homogenised in a stomacher for 6 min, transferred to a 50 mL tube, allowed to settle for 5 min and the supernatant transferred to a fresh tube. The samples were agitated on a shaker for 30 min and then incubated in an upright position for 48 h at room temperature in the dark. The supernatants were discarded and 200 µL of the pellet were transferred on each of four slopes of Herrold´s Egg Yolk Medium with Mycobactin J and Amphotericin, Nalidixic Acid and Vancomycin (HEYM, Becton Dickinson, Heidelberg, Germany). The cultures were incubated for up to six months at 37 ± 2 °C, and checked every two weeks for contamination and the occurrence of visible colonies.
Screening for Salmonella spp.
Analysis of pooled faecal samples was carried out according to ISO 6579-1:2017+A1:2020 (https://www.iso.org/standard/56712.html) after arriving in the laboratory. Buffered peptone water (BPW), modified semi-solid Rappaport-Vassiliadis (MSRV), xylose lysine deoxycholate agar (XLD) and Rambach agar (RB) were applied for the analysis (all from sifin diagnostics, Berlin, Germany).
Screening for Escherichia coli and characterization of isolates
One gram of pooled faecal material was resuspended in 9 mL of BPW (VWR International, Darmstadt, Germany). Aliquots (100 µL) of the supernatant were spread by plating onto three different solid media in Petri dishes [Gassner agar (sifin diagnostics) lacking antibiotics or supplemented with ceftiofur (4 µg/mL) or enrofloxacin (4 µg/mL)]. The plates were incubated at 37 °C for 24 h.
Antimicrobial resistant isolates
Isolates were picked from the Gassner agar plates containing ceftiofur or enrofloxacin. One colony was chosen per plate and restreaked on selective media. Species identification was performed with MALDI-TOF (Bruker, Billerica, USA) using the MALDI Biotyper Reference Library (version 3.0, Bruker). Pure cultures identified as E. coli were washed off with 2 mL LB-Miller medium containing 10% dimethyl sulfoxide (Merck, Darmstadt, Germany). One mL was stored at –80 °C and 1 mL used for preparing crude lysates by incubation at 99 °C for 15 min.
Antimicrobial sensibility testing (AST)
Isolates identified as E. coli were cultivated on Columbia-blood agar plates at 37 °C for 24 h. They were tested for antibiotic susceptibility using the Micronaut AST-system plate Micronaut-S Large Animal (MERLIN Diagnostika, Bornheim-Hersel, Germany) following the manufacturer’s instructions. In total, twelve antibiotics were tested: amoxicillin/clavulanic acid, ampicillin, ceftiofur, colistin, cephalothin, enrofloxacin, florfenicol, gentamicin, spectinomycin, trimethoprim/sulfamethoxazol, tetracycline and tulathromycin. Penicillin, erythromycin, tiamulin and tilmicosin were also present in the AST-system plate but the results were not further interpreted as E. coli are considered intrinsically resistant to these antibiotics (Moennighoff et al. 2020). Automated turbidity reading and interpretation were done according to the manufacturer’s specifications (MERLIN Diagnostika).
Detection of bla<sub>CTX-M-1</sub> group members
Members of the blaCTX-M-1 group were identified by standard PCR using previously published primers (Kim et al. 2005). The target sequence was amplified with OneTaq DNA polymerase (New England Biolabs) and 2 μL of a boiled lysate. Thermocycling conditions were as follows: 94 °C for 120 s, 25 cycles of 94 °C for 30 s, 61 °C for 60 s, and 68 °C for 45 s; and a final extension at 68 °C for 5 min. The PCR products were separated by electrophoresis for 2 h at 130 V in a 1.5% agarose gel stained with GelRed (Merck) and visualized under UV light. For Sanger sequencing, the PCR products were purified using the NucleoSpin gel and PCR clean-up kit (Macherey & Nagel, Düren, Germany) and then processed by a commercial provider (Eurofins Genomics, Ebersberg, Germany). The PCR primers were also used for sequencing and do not cover the entire reading frame of the blaCTX-M-1 group gene, so that one cannot distinguish between the blaCTX-M-1, blaCTX-M-61 and blaCTX-M-138 alleles. We therefore designated the genes as blaCTX-M-1-like.
MLVA typing
Multiple-locus variable-number tandem-repeat (VNTR) analysis (MLVA) was used for typing the E. coli isolates tested for AST (Camelena et al. 2019). It was done in two multiplex PCRs because of differences in amplification efficiency of the seven VNTR loci. The first single-tube multiplex PCR amplified five genes (rhaD, rsxC, ftsK, tolA, ytfL) using primers described previously (Camelena et al. 2019). The regions were amplified using a Taq PCR Kit (New England Biolabs, Frankfurt a. M., Germany), 2 μL of a boiled lysate, and all primers at a 10 nmol/mL concentration. Thermocycling conditions were as follows: 94 °C for 15 min, 30 cycles of 94 °C for 30 s, 58 °C for 90 s, and 68 °C for 90 s; and a final extension at 68 °C for 10 min. The second PCR amplified variable regions in the loci tRNA-Arg and hemY using primers described previously (Camelena et al. 2019). The regions were amplified by a single-tube duplex PCR using a Taq PCR Master Mix Kit (Qiagen), 2 μL of a boiled lysate, and all primers at concentrations of 10 nmol/mL. The thermocycling conditions were as follows: 95 °C for 15 min, 30 cycles of 94 °C for 30 s, 55 °C for 90 s, and 72 °C for 90 s; and a final extension at 72 °C for 10 min. The PCR-amplified fragments of both multiplex PCRs (10 μL) were separated by electrophoresis for 2 h at 130 V in a 2.5% agarose gel pre-stained with Gel red (Merck) and visualized under UV light. Each gel contained two flanking lanes loaded for normalization with a 100 bp DNA ladder (New England Biolabs).
Phylogenetic analysis using MLVA
PCR fingerprints of the agarose gel electrophoresis were visualized using SnapGene Viewer (GSL Biotech LLC, Chicago, USA). TIFF image files of the gels were loaded in BioNumerics software v. 7.6.3 (Applied Maths, Sint-Martens-Latem, Belgium). The molecular weight marker contained within each gel (100-bp DNA ladder) enabled normalization between different gel runs. Two experiment types were done, one for the gels with five amplified genes (MLVA_5) and the other for the gels with two amplified genes (MLVA_2). The same parameters were applied to both analyses. The unweighted pair group method using average linkages (UPGMA) was used for the dendrogram analysis. The similarity coefficient pairwise Dice was performed and the MLVA patterns were compared using an optimization parameter of 2% and a tolerance parameter of 1%. Both experiment types were integrated in a composite data set and the weight of the MLVA_5 was set double the weight of the MLVA_2. The similarity coefficient of the comparison was the average from both experiments and the clustering method used was UPGMA.
Detection of <em>stx1</em> and <em>stx2</em>
The Gassner agar plates with bacterial growth were used to isolate bacterial pools to detect E. coli via the uidA gene and to screen for the Shiga toxin encoding genes stx1 and stx2. The target sequences were amplified in a multiplex PCR with primers described previously (Muller et al. 2007, Shimpoh et al. 2017) at a concentration of 10 nmol/mL each in a single-tube multiplex PCR using a Taq PCR Kit (New England Biolabs) and 2 μL of a boiled lysate. Thermocycling conditions were as follows: 94 °C for 30 s, 30 cycles of 94 °C for 30 s, 56 °C for 40 s, and 68 °C for 40 s; and a final extension at 68 °C for 5 min. The PCR-amplified fragments (in a volume of 10 μL) were separated by electrophoresis for 55 min at 100 V in a 1.5% agarose gel stained with Gel red (Merck) and visualized under UV light. Aliquots of the bacterial pools positive for uidA and at least one of the stx genes were cultivated on Gassner agar plates at 37 °C for 24 h. Between 60–80 colonies per plate were screened for the presence of stx1 and/or stx2 using the multiplex PCR protocol for stx1/stx2/uidA gene detection described in this paragraph.
Whole genome sequencing
Strains were grown overnight at 37 °C in 5 mL of Luria-Bertani broth (Carl Roth GmbH, Karlsruhe, Germany). The genomic DNA was purified using the DNeasy® UltraClean® Microbial Kit (Qiagen). The concentration of the DNA was determined using a NanoDrop™ One/OneC Microvolume UV-Vis Spectrophotometer (Thermofisher, Dreieich, Germany). DNA sequencing libraries were prepared and paired-end sequencing was performed by Eurofins Genomics Europe Sequencing (Constance, Germany).
We performed bioinformatics analysis of the strains using the WGSBAC „in-house“ pipeline (https://gitlab.com/FLI_Bioinfo/WGSBAC). Illumina raw reads were subjected to quality control using FastQC v. 0.11.7 (available at http://www.bioinformatics.babraham.ac.uk/projects/fastqc/) and coverage was calculated using an adapted script (https://github.com/raymondkiu/fastq-info/blob/master/fastq_info_3.sh). Reads were de novo assembled using SPAdes (v. 3.15) (Bankevich et al. 2012) and evaluated with QUAST v. 5.0.2 (Gurevich et al. 2013) with standard settings. Annotation was performed with Prokka (v. 1.14.5) (Seemann 2014). The pipeline used Kraken 2 (v. 1.1) to identify contamination and the database Kraken2DB to classify both reads and contigs (Wood et al. 2019). Genes coding for resistance and punctual chromosomal mutations were detected using the software AMRFinderPlus (v. 3.10) (Feldgarden et al. 2020). Additionally, Abricate v. 1.0.1 (available at https://github.com/tseemann/abricate) with the databases ResFinder (v. 3.2) (Zankari et al. 2013), CARD (v. 3.0.8) (Jia et al. 2017) and NCBI were used for resistance gene detection. Abricate was also used in conjunction with the Virulence Factor Database (VFDB) (Liu et al. 2019) for the prediction of virulence-associated genes.
For genotyping, classic seven gene multilocus sequence typing (MLST) (v. 2.16.1) (Wirth et al. 2006) was used on assembled genomes with automatic scheme detection. In addition, cgMLSTs were assigned by submitting raw reads to the Center for Genomic Epidemiology website (http://www.genomicepidemiology.org/) using cgMLSTFinder 1.1 which runs KMA (Clausen et al. 2018) against a chosen core genome MLST (cgMLST) database, here for E. coli (Zhou et al. 2020).
Results
Detection of Clostridium difficile
C. difficile was isolated from composite faecal matter by enrichment cultures only. In total, eight isolates proved to be C. difficile positive by cdd3-targeted PCR (Table 1), resulting in 8.5% (n = 8/94, 95% CI 2.8–14.1%) positive samples and a prevalence of 16.3% (n = 7/43, 95% CI 5.2–27.3%) at farm-level.
Detection of Chlamydia
Chlamydia DNA was present in all three types of samples (Supplementary Material 3). Of the 94 faecal samples collected in 43 holdings, seven samples (7.4%, 95% CI 2.1–12.7%) from six holdings (14.0%, 95% CI 3.6–24.3%) presented as Chlamydiaceae-positive (Table 1). Two of these samples tested positive for C. pecorum by species-specific qPCR, while differentiation of the other samples was not possible due to low Chlamydia DNA copy numbers. Six out of 136 vaginal swabs (4.4%, 95% CI 0.9–7.8%) from four of 24 holdings (16.7%, 95% CI 1.7–31.6%) were Chlamydia-positive (Supplementary Material 3). Species identification was not successful due to low DNA concentrations. In one of twelve placental samples (8.3%) from one of nine holdings (11.1%), Chlamydiaceae were detected and identified as C. pecorum (Supplementary Material 3). There were no correlations between positive faecal samples and positive vaginal swab samples in terms of farm origin. However, the positive placenta sample originated from a farm with Chlamydia-positive faecal sample results (farm 10) (Supplementary Material 3). The overall concentration of Chlamydia DNA was low with Ct values ranging between 33.0 and 37.9 (Table 1).
Detection of Mycobacterium avium subsp. paratuberculosis (MAP) and Salmonella spp.
Neither cultural growth of MAP nor of Salmonella organisms was detected in any of the faecal samples examined (Supplementary Material 3).
Detection and characterization of Shiga toxin-producing Escherichia coli
In total, 18% of the faecal samples (n = 17/94) (95% CI 10.3–25.9%) originating from twelve holdings (28%; 95% CI 14.5%–41.3%), were positive for stx1 and/or stx2 (Table 1 and 2). The stx genes were usually detected in the pool washed off the Gassner agar plate without antibiotics. Only one stx1 positive sample was derived from a Gassner agar plate containing enrofloxacin (19SC0267) (Table 2). In total, twelve of the samples contained stx1 and one stx2, and both genes were found in four faecal samples (Table 1 and 2).
Six strains were isolated from these samples, three carried stx1, two stx2 and one both genes. The samples originated from four holdings, and in two of them (holdings #6 and #31) two isolates with different stx genes were detected (Table 2). The six STEC isolates were assigned to different serogroups and the O antigen of one isolate (19BP0191-3-D-8) (Table 3) was not typeable with the SerotypeFinder tool, but could be assigned to OX18 (DebRoy et al. 2016) (C. Lang and A. Fruth, personal communication). None of the strains belonged to the O157 or the “Big Six” non-O157 serogroups (Table 3). Four isolates belonged to phylogroup B1 and two (19BP0127-1-A and 19BP0332-13-56) were classified as phylogroup A. These two strains belonged to the same ST (2101) and cgMLST (15459), and the other isolates were assigned different STs (Table 3). Antimicrobial resistance genes (ARGs) were found in only one isolate, 19BP0191-3-D-8, which carried six ARGs (aadA5, blaTEM-1B, dfrA17, fosA7, mph.A, sul1) and three chromosomal mutations mediating high-level fluoroquinolone resistance (gyrA_D87N, gyrA_S83L, parC_S80I) (Table 3). The stx1 and/or stx2 gene presence corresponded to the PCR-amplified fragments detected; however, in one isolate’s genome sequence (19BP0399-5-25), no stx gene was found (Table 3). The adherence factor gene ompA was detected in all isolates and the cluster encoding fimbrial adhesins (AFA) was found in one isolate (19BP0271). The intimin encoding gene eae was not detected in any strain (Table 3).
Detection and characterization of antimicrobial-resistant Escherichia coli
Primary screening for the presence of E. coli was done with the bacterial culture isolated from the Gassner plate without antibiotics using primers for the E. coli-specific gene uidA (Shimpoh et al. 2017). Positive signals indicating E. coli were present in 93 of 94 faecal samples (98.9%) representing all farms (100%) (Supplementary Material 3). A total of 148 colonies was isolated, 85 colonies from Gassner agar plates with ceftiofur and 63 from Gassner agar plates with enrofloxacin (4 µg/mL). One bacterial colony was taken per plate; colonies were chosen primarily based on their blue colour (lactose positive), but green to yellow colonies were also picked in the absence of lactose-positive colonies. MALDI-TOF identified 42.7% of these isolates (63/148) as E. coli. The remaining belonged to other genera; the most frequently identified were Acinetobacter spp., Pseudomonas spp. and Providencia spp. (Supplementary Material 4).
All 63 E. coli isolates, obtained from faecal samples from 25 holdings (58.1%), were resistant to at least five antibiotics and three different antibiotic classes (Table 4). High levels of non-susceptibility (i.e., classified as resistant or of intermediate susceptibility) were found against ampicillin (85.7%) and cephalothin (95%). More than half of the isolates (57%) were non-susceptible to enrofloxacin, 43% to ceftiofur, which almost corresponds to the numbers of strains obtained from the respective selective media, except for five isolates, which were non-susceptible to both antibiotics. Possible ESBL producers were found in 31% (n = 29/94) (95% CI 21.7%–40.3%) of the faecal samples and in 46% (n = 29/63) of the total isolates (Table 4). ESBL enzymes of the CTX-M-1 group (Bonnet 2004) are frequently found in humans, companion animals and livestock from Europe (Ewers et al. 2012). The ceftiofur-resistant strains were, therefore, probed for CTX-M β-lactamase presence using a blaCTX-M-1 group specific PCR (Kim et al. 2005). Twenty-six out of 30 strains were PCR-positive and ten strains arbitrarily selected for Sanger sequencing classified as CTX-M-1-like, confirming the ESBL phenotype (Supplementary Material 5). In contrast, a high percentage of the isolates retained susceptibility to gentamicin (97%), amoxicillin/clavulanic acid (97%), tulathromycin (89%) and colistin (83%) (Table 2).
Among the E. coli isolates, 43 different MLVA profiles were recorded (Fig. 2). The profiles found formed two big clusters with the first containing 34 MLVA profiles and the second nine MLVA profiles. Thirty isolates produced a unique pattern and thirteen MLVA types were shared between more than one isolate (Fig. 2). Seven of the latter were detected in isolates belonging to different holdings. Five strains with identical MLVA type differed in their antimicrobial resistance patterns (Supplementary Material 6).
Questionnaire metadata
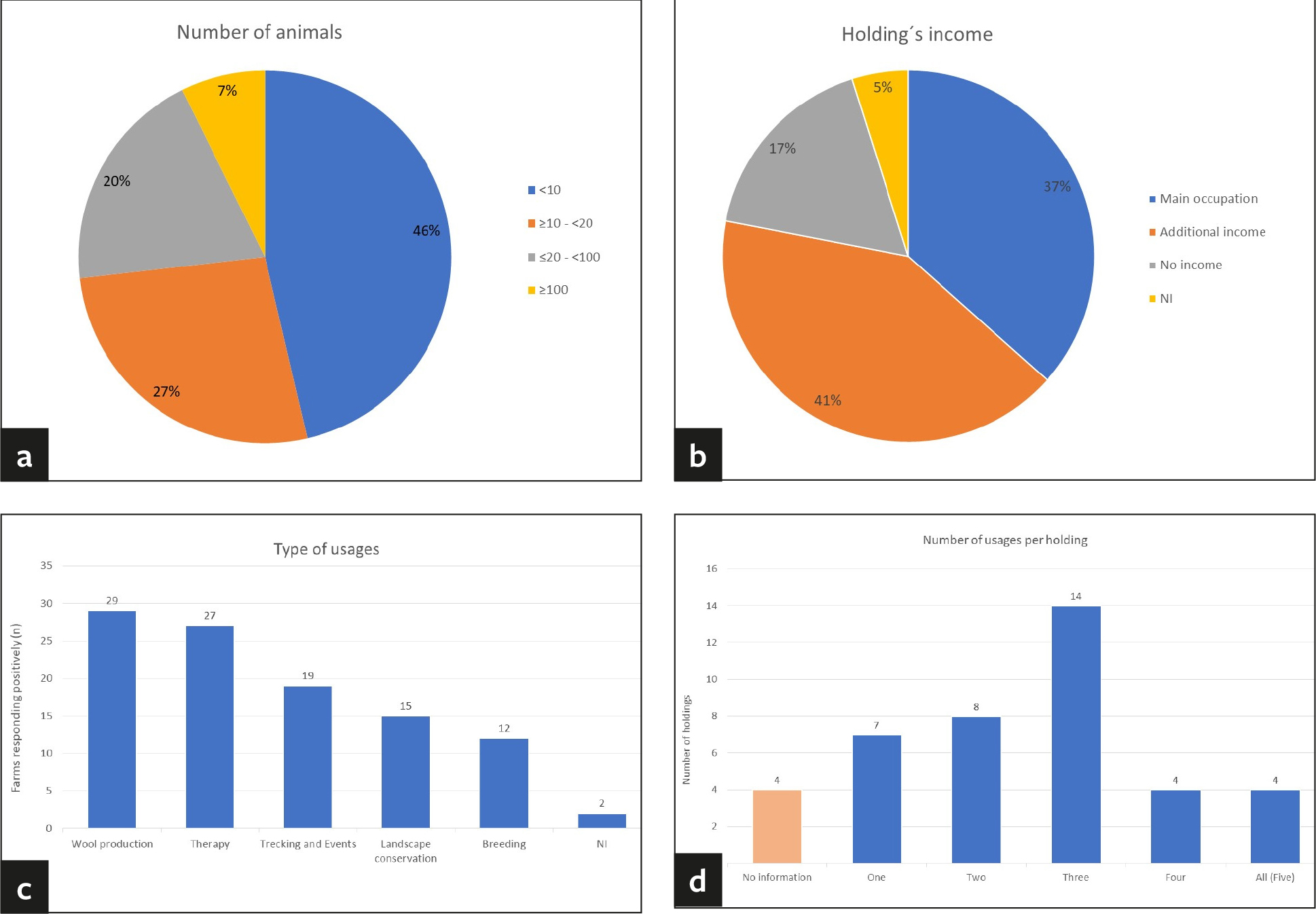
Of the 43 holdings sampled, most were located in Saxony (63%, n = 27/43), followed by Thuringia (21%, n = 9/43) and Saxony-Anhalt (16%, n = 7/43) (Supplementary Material 1). The questionnaire was partially or fully completed by 41 holdings, representing a total of 974 animals. The majority of the animals were alpacas and only 2.1% (n = 20/974) were llamas. Most of the farms were smallholdings (Fig. 3a). Almost half housed fewer than ten animals (46%, n = 19/41) and 27% of the farms (n = 11/41) kept between ten and 20 animals. A further 20% of the farms (n = 8/41) kept between 20 and 100 animals and only 7% (n = 3/41) were large holdings with more than 100 animals (Fig. 3a). Approximately one third (37%, n = 15/41) of the holdings kept SAC as their principal occupation and 42% (n = 17/41) as source of additional income (Fig. 3b). Only 17% (n = 7/41) raised the animals without generating income and two farms did not respond to this particular question (Fig. 3b). The most common type of usage on farms was wool-production followed by therapy, trekking, landscape conservation and breeding (Fig. 3c). Usually, the farms mentioned more than one activity, only 17% (n = 7/41) kept SAC for one specific type of usage only (Fig. 3d). Of all herds, 22% (n = 9/41) were supplemented in the recent past by purchase of animals from other sources while the remaining owners claimed to follow a closed herd concept. It must be noted, though, that only 61% (n = 25/41) of the owners replied to that question in the questionnaire. In most instances, 51% (n = 21/41) of the holdings, the animals purchased were raised in Germany. Five out of 41 owners each (12%) bought animals from the EU member states and/or from other parts of the world, such as Australia and North and South America. Frequently, animals belonging to other species than SAC were housed on the same premises; 49% (n = 20/41) of the farms also kept ruminants and other domestic animals and 24% (n = 10/41) were zoos with multiple animal species. Of all owners, 15% (n = 6/41) indicated that the SAC are in direct contact with other animals (ducks, pony, and sheep). Persons in contact with SAC were usually the owners, the veterinarian, and the working staff. However, 76% (n = 31/41) of the farms frequently welcomed visitors. Staff and/or visitors had contact with SAC and other animals on 83% (n = 34/41) of the holdings.
Discussion
The number of SAC kept in Germany has increased during the last decades. They are frequently in close contact with other animals and humans. This study aimed at improving our knowledge on their health status and the potential to act as a source of transmission of infectious agents. In fact, zoonotic and pathogenic bacteria and multiresistant indicator E. coli were detected in SAC in Central Germany.
C. difficile is widely distributed in the environment, is found occasionally in faecal samples of multiple species, and is known for its zoonotic potential (Diaz et al. 2018). The fact that isolates were obtained only after enrichment culture suggests a relatively low abundance of C. difficile organisms in the SAC faeces (Table 1). A detailed characterization of the isolates obtained will be presented in a follow-up study. To our knowledge, there are no reports of C. difficile as a cause of enteritis in llamas and alpacas. However, C. difficile can cause enteritis in some animal species, e.g. horses and pigs, while its role in disease in other species, like e.g. cattle, dogs and poultry, is not known (Diab et al. 2016). Therefore, testing for C. difficile should be considered as a possible differential diagnosis for intestinal disease in llamas and alpacas. Further studies on the prevalence of C. difficile and the types present in llamas and alpacas should be conducted to assess the risk of zoonotic transmission.
Veterinarians frequently associate keratoconjunctivitis, arthritis and reproductive disorders in SAC (abortions, stillbirths, and births of weak crias) with chlamydial agents. Considering the high loss rate of crias in domesticated alpaca and llama herds (Neubert et al. 2021), and the fact that C. abortus is held responsible for about 45% of infectious abortions and dead/weak lambs in small ruminants (Essig and Longbottom 2015), it is surprising that no systematic epidemiological or experimental investigations involving this pathogen have been conducted in SAC so far. There are only a few reports on the presence of chlamydiae or Chlamydia-specific antibodies in SAC (Probst 2008, Li et al. 2020) and no proof of an etiological role. Interestingly, vaccination against C. abortus with vaccines approved for small ruminants is recommended even though no studies on the effectiveness in SAC are available (Neubert et al. 2021). In sheep, both C. abortus and C. pecorum are endemic in Europe and are transmitted via the oronasal route. While C. pecorum establishes an asymptomatic infection in the gastrointestinal tract or rarely causes arthritis, C. abortus is latent in rams and non-pregnant ewes, but recrudesces and initiates placentitis and late abortion in pregnant ewes (Essig and Longbottom 2015). The most important source of transmission to naïve ewes and to humans, where the infection with C. abortus can cause severe systemic disease and abortion, results from the products of abortion. Extremely large numbers of chlamydiae can be detected in the vaginal discharges, placentas and on the coats of fetuses after abortion or parturition (Essig and Longbottom 2015). Intermittent weak shedding occurs in faeces and vaginal discharges independent of the reproductive processes, which can indicate the presence of the infectious agents in a herd (Lenzko et al. 2011). The findings of this study imply that chlamydiae are present in German SAC herds, but the low chlamydial load in the samples hindered genotyping to the species level in most instances. Although only C. pecorum was identified in faecal and placental samples, the presence of C. abortus cannot be excluded (Table 1, Supplementary Material 3). No cases of acute chlamydiosis were detected in this study. To unequivocally unveil the role of chlamydiae in reproductive disorders of SAC and to assess a potential zoonotic risk, tissue samples from abortion material and vaginal swab samples from mares should be collected shortly after abortion or parturition. In addition, serological examination would be helpful to estimate the prevalence of chlamydial infection, especially for C. abortus at the herd level. However, such surveys are hampered by the lack of validated serological tests.
Few studies have been conducted to investigate the occurrence of Mycobacterium avium subsp. paratuberculosis (MAP) in SAC, either in their natural habitats in the Andes of South America, or in the countries where they have been exported to. MAP infections do affect SAC, but in general, the prevalence of Paratuberculosis seems to be rather low in these animal species. MAP was isolated from the faeces of 4.2% of wild guanacos on Tierra del Fuego Island and from 15/85 free-ranging alpacas inhabiting the Chilean Altiplano (Salgado et al. 2009, 2016). No antibodies against MAP were found in 207 free-ranging and captive vicuñas and 614 domestic camelids (alpacas and llamas) in a natural reserve in Peru (Risco-Castillo et al. 2014). Outside of South America, mainly individual case reports of Paratuberculosis in llamas and alpacas have been published (Belknap et al. 1994, Fecteau et al. 2009, Munster et al. 2013). This corresponds with the low prevalence observed, but may also be due to the fact, that the disease will only hardly be suspected following clinical observation. Like cattle, SAC infected with MAP develop a granulomatous enteritis in the small intestines, leading to protein-losing enteropathy and severe weight loss. However, SAC often do not develop the intractable diarrhoea that accompanies the weight loss in cattle. Any weight loss is often obscured by their fleece (Fecteau et al. 2013) and the disease might, therefore, be easily missed by veterinarians. The present study is the first to address the prevalence of MAP infection in domestic SAC in Germany. The negative results indicate that MAP is not present or only present at a very low prevalence in the SAC herds in Central Germany. These findings are in line with data from Austria, where MAP could not be culturally isolated from the faeces of 184 individual llamas and 261 alpacas on 78 farms, while 7.2% of the llama samples and 1.6% of the alpaca samples were positive using qPCR (Stanitznig et al. 2017) which was not applied in our study. A survey on MAP faecal shedding in alpacas presented to veterinary hospitals in the United States revealed a prevalence of 6% faecal shedders (Fecteau et al. 2013). This prevalence must be treated with caution because the study population was pre-selected for health issues and, therefore, not entirely representative for the national alpaca herd in general.
Salmonella (S.) organisms represent one of the most important zoonotic agents as well as a cause of subclinical and clinical disease in numerous animal species. Compared to ruminants, little is known about the significance or prevalence of Salmonella spp. in SAC (Staples 2016). The detection of twenty-three infections caused by seven Salmonella serovars, including two abortions by S. Dublin and S. Typhimurium between 2000–2015 in England and Wales (Halsby et al. 2017) indicate both, the basic occurrence of different Salmonella serovars in SAC with or without clinical manifestation but also a rather low prevalence in these species. Examination of 94 composite faecal samples from 43 holdings located in three federal states in this study did not yield any Salmonella detection, pointing to a very low prevalence in the region under study.
STEC cause severe human food- and waterborne infections worldwide. The major STEC reservoir is the gastrointestinal tract of ruminants (Gyles 2007), but STEC were found to be shed by many other animal species (Persad and LeJeune 2014) including SAC, e.g., in Peru and the United Kingdom (Mercado et al. 2004, Silvera et al. 2012, Halsby et al. 2017, Maturrano et al. 2018, Siuce et al. 2020). This study has isolated the first STEC from German alpacas. Composite faecal samples taken from 17 groups of animals on twelve farms were positive for stx1 and/or stx2, representing 18.1% of all 94 faecal samples taken (Table 1 and 2). The strains isolated belonged to different non-O157 serogroups (Table 3), but the serogroups identified have been associated with human clinical cases of diarrhoea (O55) (Valilis et al. 2018), or were also isolated from other animals, such as horses (O87) (Golomidova et al. 2021), sheep (O76) (Michelacci et al. 2013) and wild birds (O86) (Feng et al. 2005). For one of the isolates, an O antigen was not assigned bioinformatically (Table 3) by the SerotypeFinder tool, which is not unusual, because 8% of E. coli isolates have no O antigen (Gyles 2007). The presence of the stx1 and stx2 genes was confirmed in the WGS analysis (Table 3). Four strains carried stx1 or stx2, and one carried both genes. In one of the isolates, no stx gene was found, despite its presence in prior PCR analysis (Table 2 and 3). The loss of an stx-encoding gene or the stx-encoding phage is not uncommon in E. coli (Barth et al. 2016). An intimin-encoding eae gene, believed to be indicative of a higher risk of the respective strain to cause severe human illness (Gyles 2007, Scheutz 2014, FAO 2019), was absent in the strains obtained. However, the adhesin gene ompA was found in all isolates and one isolate encoded the Afa/Dr cluster of fimbrial adhesins, expressed by uropathogenic and diffusely adhering E. coli (Table 3). Much enforced by the occurrence of a large STEC outbreak in Germany in 2011, caused by an unusual O104:H4 strain, which was eae-negative but possessed other means of bacterial intestinal adherence, EFSA (EFSA et al. 2020) nowadays classifies all STEC strains as potential human pathogens. Detection of STEC in German SAC, therefore, needs to be regarded as a potential zoonotic threat.
SAC are subjected to antibiotic treatment, which is a major driver of antimicrobial resistance in bacteria (Shallcross and Davies 2014). Increasing spread of AMR bacteria documented for several animal species in recent times has raised much concern, both from a veterinary clinical standpoint but also from a One Health perspective. Different from other livestock species, neither antimicrobial usage nor occurrence and prevalence of AMR bacteria in SAC are currently systematically assessed and recorded in Germany. Analysis of the E. coli population in the faecal matter from SAC was therefore used as a surrogate to estimate the presence of AMR in these animals. Our results showed that all isolates were at least resistant against five antibiotics and three different antibiotic classes (Table 4, Supplementary Material 6). In addition, 43% (n = 29/63) of the isolates were proposed to be ESBL producers and 26 of these isolates were PCR-positive for members of the blaCTX-M-1 group (Supplementary Material 3, Supplementary Material 5). Since there are many different ESBL β-lactamases (Bush 2018), the remaining PCR-negative, ceftiofur-resistant strains will be characterized later by WGS. The high percentage of CTX-M-1 group enzymes agrees with their frequent identification in resistant strains isolated from animals in Europe (Ewers et al. 2012) (Table 3, Supplementary Material 6). A high degree of non-susceptibility was found against ampicillin and cephalothin. Resistance to enrofloxacin and ceftiofur was high, reflecting the initial selection step, but several of the isolates were also resistant to both antibiotics (Table 3, Supplementary Material 6). Some information about resistance against anthelmintic drugs (Galvan et al. 2012) and about bacteria involved in dental abscessation in SAC (Niehaus and Anderson 2007), where different Gram-negative bacteria, including E. coli, were present, is available. As a group, the bacterial species were resistant against neomycin (33%, n = 32/98), enrofloxacin (19%, n = 19/98), gentamicin (17%, n = 17/98), sulfisoxazole (13%, n = 13/98), but all remained susceptible to ceftiofur (Niehaus and Anderson 2007). In Peru, two studies demonstrated for E. coli strains intermediate to high levels of resistance against antibiotics. Most of the E. coli strains were resistant to neomycin (n = 41/51) and some of them to oxytetracycline (n = 13/51) (Luna et al. 2012). In another study, most of the isolates from alpacas with or without diarrhea displayed resistance to nitrofurantoin (n = 69/82), and a low number of isolates were resistant to amikacin (n = 10/82), ceftazidime (n = 9/82) or enrofloxacin (n = 9/82) (Barrios-Arpi et al. 2016). In Germany, the prevalence of ESBL/AmpC E. coli from selective sampling determined for fattening calves and for young cattle at the slaughterhouse was 70.8% (n = 288/407) in 2019 (BVL 2020). In fattening pigs and in pigs at the slaughterhouse, 39.6% (n = 152/384) and 49.1% (n = 192/391) of the colonic selective samples carried ESBL/AmpC expressing isolates, respectively (BVL 2020). CTX-M-1 positive E. coli are usually found in animals, mostly in horses, chicken and swine (Cormier et al. 2019); our study shows the first detection of blaCTX-M-1 in SAC. The results imply that the AMR situation in Central German SAC appears to be similar to that in other livestock animal species, suggesting that the same set of rules and regulations should be adopted for AMR management in SAC.
The SAC head counts have increased in Europe and other continents during the last decade (D’Alterio et al. 2006, Bauerstatter et al. 2018, Zarrin et al. 2020, Neubert et al. 2021) and appear to continue to increase. It is, therefore, not only important to obtain information about the diseases affecting these animals but also their breeding and holding conditions, as the latter may affect the former. Several studies with questionnaires have been done in the UK (farms n = 86, SAC n = 462) (D’Alterio et al. 2006), Austria (farms n = 218, SAC n = 3520) (Bauerstatter et al. 2018) and Germany (farms n = 220, SAC n = 3803) (Neubert et al. 2021) for gathering information, among others, on SAC, their breeding, husbandry, and medical treatment. In our study, 974 animals were kept in the 41 holdings that returned the questionnaire. Most kept alpacas, only 2% (n = 20/974) of the animals were llamas. In the Austrian study, 39% (n = 34/86) of the farms kept llamas (Bauerstatter et al. 2018). Lower numbers of llamas were reported for the UK (D’Alterio et al. 2006) and Germany (Neubert et al. 2021) with 20.6% (n = 726/2719) and 10.6% (n = 27/255), respectively. Most of the owners have smallholdings (75%, n = 30/41) with less than 20 animals (Fig. 3a), similar to the UK with 65% (n = 142/218) (D’Alterio et al. 2006) and Austria with 77% (n = 66/86) (Bauerstatter et al. 2018). Most of the SAC farms previously surveyed in Germany were located in North Rhine-Westphalia, Lower Saxony and Bavaria (Neubert et al. 2021). Our study approached farms in Central-Germany and adds to the dataset, although data from several parts of Germany is still missing. One third of the owners keep SAC as their main occupation (Fig. 3b), in contrast to 8.2% (n = 21/255) as determined by Neubert et al. (2021), indicating regional differences. Wool-production is the most common intended use followed by therapy, trekking, landscape conservation and breeding (Fig. 3c). Other studies reported similarly, although the hobby aspect was more prominent (D’Alterio et al. 2006, Bauerstatter et al. 2018, Neubert et al. 2021). Almost half of the owners keep other animal species, such as domestic animals and ruminants, in addition; again, like the other studies (D’Alterio et al. 2006, Bauerstatter et al. 2018, Neubert et al. 2021). However, only 15% (n = 6/41) of the holdings in this study mentioned direct contact of their SAC with other animals mirroring the low percentage (6.7%, n = 17/255) described by Neubert et al. (2021) but clearly different from the 28% (n = 59/218) reported for the UK (D’Alterio et al. 2006). In our study, the SAC usually are in contact with the owners, the staff and the veterinarians. Remarkably, 75% (n = 31/41) of the holdings frequently receive visitors and 83% (n = 34/41) of the staff and/or the visitors have contact with the SAC and the other animals. Unfortunately, the other studies did not provide this type of information, despite its importance because contact can act as a vector from SAC to other animals or to humans, thereby promoting the transmission of AMR and zoonotic bacteria. Only approximately one quarter of the owners (22%, n = 9/41) stated that they had bought SAC in recent years, 61% (n = 25/41) disclosed the source for recruiting animals to be Germany. Animals originating from the EU and other parts of the world, such as Australia and Nord and South America were purchased by similar percentages of owners (12.2%, n = 5/41 each). The data from Neubert et al. (2021) shows a similar tendency with 12% of the owners (n = 30/255) having animals originating from diffent flocks in Germany and only 3.5% (n = 9/255) with animals from other EU or non-European countries. In 2006, it was published for the UK that 45% (n = 97/216) of the owners imported animals from one or more South American countries, most frequently from Chile (D’Alterio et al. 2006). Taking into account all the data and the fact that the number of animals in the EU has increased strongly during the last decade, a frequent trade of animals between holdings and across borders within and from outside the EU must occur and should not be underrated from an epidemiological viewpoint. Of note, these animal movements presumably occurred with little or no testing for infectious diseases prior to the novel European Animal Health law coming into effect.
Conclusion
Our study results indicate that zoonotic and pathogenic bacteria and multiresistant indicator E. coli are present in SAC in Central Germany. Because of possible regional differences in SAC farming conditions, further studies are required for conclusions on the real prevalence or on risk factors for the occurrence of the pathogens detected and not detected in the present study. Results from this study do not exclude that close contact to other farm animals (sheep, cattle, goats) or sharing of pastures with other animals might be a source of Salmonella or MAP infections in SAC. The potential risk of SAC-derived transmission of epizootic and zoonotic pathogens to other animals including livestock and to humans seems to be low. However, future studies have to systematially complete the dataset to cover all parts of Germany and include additional bacterial (e.g. Mycobacterium tuberculosis, Coxiella burnetii) and viral (e.g. Orthopox Virus) agents before a comprehensive picture of the heath status of and the zoonotic risk potential associated with SAC in Germany is gained which will be needed for establishing monitoring and preventive medicine programs to address health threats to and from SAC in Germany.
Acknowledgments
We thank Dr. Elisabeth Wiegand for her contributions to study design, holding recruitment and sampling. Dr. Bodo Kröll is acknowledged for providing diagnostic samples and background data which were included in this study. We thank Kerstin Barth, Simone Bettermann, Annegret Jahn, Silke Keiling, Peggy Methner, Maike Richter, Sandy Werner for skillful technical assistance.
Ethical approval
All relevant international, national and/or institutional ethical guidelines for the handling and care of the animals used in the study were observed. The information regarding the application for approval of animal experimentation and its authorization is given in the publication.
Conflict of interest
The authors hereby declare that they have no proprietary, professional or other personal interests in any product, service and/or company that could have influenced the contents or opinions expressed in this publication.
Funding
For this work, Belén González-Santamarina was supported by the joint research project JRP13-AMRSH5-WORLDCOM, which has received funding from the European Union’s Horizon 2020 research and innovation program under Grant Agreement No 773830.
Data Availability Statement
The raw sequence data generated during the current study is available at https://www.ncbi.nlm.nih.gov/bioproject/PRJNA776159. The respective accession numbers of the individual strains’ sequences can be found in Table 3.
Authors contribution
Conception or design of the project: BGS, CB, CM.
Experimental analysis and interpretation: BGS, CSc, CSe, HK, MW, UM.
Drafting of the manuscript: BGS, CB, CSc, CSe, HK, MW, UM, CM.
All authors approved the final version.
Address for correspondence
Belén González-Santamarina
Institute of Molecular Pathogenesis (IMP)
Friedrich-Loeffler-Institut
Naumburger Str. 96a
07743 Jena
Germany
belengsan@gmail.com
References
Bankevich A, Nurk S, Antipov D, Gurevich AA, Dvorkin M, Kulikov AS, Lesin VM, Nikolenko SI, Pham S, Prjibelski AD, Pyshkin AV, Sirotkin AV, Vyahhi N, Tesler G, Alekseyev MA, Pevzner PA (2012): SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J Comput Biol 19(5): 455–477.
Barrios-Arpi M, Siever M. C, Villacaqui-Ayllon E (2016): Susceptibilidad Antibiótica de Cepas de Escherichia coli en Crías de Alpaca con y sin Diarrea. Rev Invest Vet Perú 27: 381.
Barth SA, Menge C, Eichhorn I, Semmler T, Wieler LH, Pickard D, Belka A, Berens C, Geue L (2016): The Accessory Genome of Shiga Toxin-Producing Escherichia coli Defines a Persistent Colonization Type in Cattle. Appl Environ Microbiol 82(17): 5455–5464.
Bauerstatter S, Lambacher B, Stanitznig A, Franz S, Wittek T (2018): Neuweltkamele in Österreich – Untersuchungen zur Population, Haltung, Herdenmanagement und Gesundheitsprophylaxe. Wien Tierärztl Monatsschrift 105: 191–199.
Belknap EB, Getzy DM, Johnson LW, Ellis RP, Thompson GL, Shulaw WP (1994): Mycobacterium paratuberculosis Infection in 2 Llamas. J Am Vet Med Assoc 204(11): 1805–1808.
Blankenship HM, Mosci RE, Dietrich S, Burgess E, Wholehan J, McWilliams K, Pietrzen K, Benko S, Gatesy T, Rudrik JT, Soehnlen M, Manning SD (2021): Population structure and genetic diversity of non-O157 Shiga toxin-producing Escherichia coli (STEC) clinical isolates from Michigan. Sci Rep 11(1): 4461.
Bonnet R (2004): Growing group of extended-spectrum beta-lactamases: the CTX-M enzymes. Antimicrob Agents Chemother 48(1): 1–14.
Bush K (2018): Past and Present Perspectives on beta-Lactamases. Antimicrob Agents Ch 62(10): e01076-18.
BVL (2020): Berichte zur Lebensmittelsicherheit 2019 – Zoonosen-Monitoring. www.bvl.bund.de/ZoonosenMonitoring (21.06.2021).
Camelena F, Birgy A, Smail Y, Courroux C, Mariani-Kurkdjian P, Le Hello S, Bonacorsi S, Bidet P (2019): Rapid and Simple Universal Escherichia coli Genotyping Method Based on Multiple-Locus Variable-Number Tandem-Repeat Analysis Using Single-Tube Multiplex PCR and Standard Gel Electrophoresis. Appl Environ Microbiol 85(6): e02812-02818.
Clausen P, Aarestrup FM, Lund O (2018): Rapid and precise alignment of raw reads against redundant databases with KMA. BMC Bioinformatics 19(1): 307.
Cormier A, Zhang PLC, Chalmers G, Weese JS, Deckert A, Mulvey M, McAllister T, Boerlin P (2019): Diversity of CTX-M-positive Escherichia coli recovered from animals in Canada. Vet Microbiol 231: 71–75.
D’Alterio GL, Knowles TG, Eknaes EI, Loevland IE, Foster AP (2006): Postal survey of the population of South American camelids in the United Kingdom in 2000/01. Vet Rec 158(3): 86–90.
DebRoy C, Fratamico PM, Yan XH, Baranzoni G, Liu YH, Needleman DS, Tebbs R, O‘Connell CD, Allred A, Swimley M, Mwangi M, Kapur V, Garay JAR, Roberts EL, Katani R (2016): Comparison of O-Antigen Gene Clusters of All O-Serogroups of Escherichia coli and Proposal for Adopting a New Nomenclature for O-Typing. Plos One 11(4): e0147434.
Diab SS, Uzal FA, Songer JG (2016): Diseases produced by Clostridium difficile. In: Uzal FA, Songer JG, Prescott J, Popoff M (eds.), Clostridial diseases of animals. Willey Blackwell, Ames, IA, 177–195.
Diaz CR, Seyboldt C, Rupnik M (2018): Non-human C. difficile Reservoirs and Sources: Animals, Food, Environment. Adv Exp Med Biol 1050: 227–243.
EFSA, ECDC (2020): The European Union Summary Report on Antimicrobial Resistance in zoonotic and indicator bacteria from humans, animals and food in 2017/2018. EFSA J 18(3): 166.
EFSA, Koutsoumanis K, Allende A, Alvarez-Ordóñez A, Bover-Cid S, Chemaly M, Davies R, De Cesare A, Herman L, Hilbert F, Lindqvist R, Nauta M, Peixe L, Ru G, Simmons M, Skandamis P, Suffredini E, Jenkins C, Monteiro Pires S, Morabito S, Niskanen T, Scheutz F, da Silva Felício MT, Messens W, Bolton D (2020): Pathogenicity assessment of Shiga toxin-producing Escherichia coli (STEC) and the public health risk posed by contamination of food with STEC. EFSA J 18(1): 5967.
Ehricht R, Slickers P, Goellner S, Hotzel H, Sachse K (2006): Optimized DNA microarray assay allows detection and genotyping of single PCR-amplifiable target copies. Mol Cell Probe 20(1): 60–63.
Essig A, Longbottom D (2015): Chlamydia abortus: New Aspects of Infectious Abortion in Sheep and Potential Risk for Pregnant Women. Curr Clin Micro Rpt 2(1): 22–34.
Ewers C, Bethe A, Semmler T, Guenther S, Wieler LH (2012): Extended-spectrum beta-lactamase-producing and AmpC-producing Escherichia coli from livestock and companion animals, and their putative impact on public health: a global perspective. Clin Microbiol Infect 18(7): 646–655.
FAO (2019): Hazard Identification and Characterization: Criteria for Categorizing Shiga Toxin-Producing Escherichia coli on a Risk Basis. J Food Protect 82(1): 7–21.
Fecteau ME, Ross J, Tennent-Brown BS, Habecker PL, Sreevatsan S, Sweeney RW, Whitlock RH (2009): Mycobacterium avium ssp. paratuberculosis. High Shedding in an Adult Female Alpaca, and its Implications for the Rest of the Herd. J Vet Intern Med 23(6): 1311–1314.
Fecteau ME, Bedenice D, Cebra CK, Pinn TL, McAdams SC, Fyock TL, Whitlock RH, Sweeney RW (2013): Prevalence of Mycobacterium avium subsp. paratuberculosis. Fecal Shedding in Alpacas Presented to Veterinary Hospitals in the United States. J Vet Intern Med 27(5): 1228–1233.
Feldgarden M, Brover V, Haft DH, Prasad AB, Slotta DJ, Tolstoy I, Tyson GH, Zhao SH, Hsu CH, McDermott PF, Tadesse DA, Morales C, Simmons M, Tillman G, Wasilenko J, Folster JP, Klimke W (2020): Validating the AMRFinder Tool and Resistance Gene Database by Using Antimicrobial Resistance Genotype-Phenotype Correlations in a Collection of Isolates. Antimicrob Agents Chemother 64(4): e00483–00419.
Feng L, Han WQ, Wang Q, Bastin DA, Wang L (2005): Characterization of Escherichia coli 086 O-antigen gene cluster and identification of O86-specific genes. Vet Microbiol 106(3-4): 241–248.
Fowler ME (2009): Camelids Are Not Ruminants. Zoo and Wild Animal Medicine 2008: 375–385. doi: 10.1016/B978-141604047-7.50049-X.
Galvan N, Middleton JR, Nagy DW, Schultz LG, Schaeffer JW (2012): Anthelmintic resistance in a herd of alpacas (Vicugna pacos). Can Vet J 53(12): 1310–1313.
Gauly M (2019): Haltung. In: Gauly M, Vaughan J, Cebra C (Hrsg.), Neuweltkameliden. 4., akt. erw. Aufl. Thieme, Stuttgart.
Gestrich A, Bedenice D, Ceresia M, Zaghloul I (2018): Pharmacokinetics of intravenous gentamicin in healthy young-adult compared to aged alpacas. J Vet Pharmacol Ther 41(4): 581–587.
Gidlewski T, Cheville NF, Rhyan JC, Miller LD, Gilsdorf MJ (2000): Experimental Brucella abortus induced abortion in a llama: pathologic effects. Vet Pathol 37(1): 77-82.
Golomidova AK, Efimov AD, Kulikov EE, Kuznetsov AS, Belalov IS, Letarov AV (2021): O antigen restricts lysogenization of non-O157 Escherichia coli strains by Stx-converting bacteriophage phi24B. Sci Rep 11(1): 3035.
Gurevich A, Saveliev V, Vyahhi N, Tesler G (2013): QUAST: quality assessment tool for genome assemblies. Bioinformatics 29(8): 1072–1075.
Gyles CL (2007): Shiga toxin-producing Escherichia coli: an overview. J Anim Sci 85(13 Suppl): E45–62.
Halsby K, Twomey DF, Featherstone C, Foster A, Walsh A, Hewitt K, Morgan D (2017): Zoonotic diseases in South American camelids in England and Wales. Epidemiol Infect 145(5): 1037–1043.
Hoffmann B, Depner K, Schirrmeier H, Beer M (2006): A universal heterologous internal control system for duplex real-time RT-PCR assays used in a detection system for pestiviruses. J Virol Methods 136(1-2): 200–209.
Jia BF, Raphenya AR, Alcock B, Waglechner N, Guo PY, Tsang KK, Lago BA, Dave BM, Pereira S, Sharma AN, Doshi S, Courtot M, Lo R, Williams LE, Frye JG, Elsayegh T, Sardar D, Westman EL, Pawlowski AC, Johnson TA, Brinkman FSL, Wright GD, McArthur AG (2017): CARD 2017: expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res 45(D1): D566–D573.
Kim J, Lim YM, Rheem I, Lee Y, Lee JC, Seol SY, Lee YC, Cho DT (2005): CTX-M and SHV-12 beta-lactamases are the most common extended-spectrum enzymes in clinical isolates of Escherichia coli and Klebsiella pneumoniae collected from 3 university hospitals within Korea. FEMS Microbiol Lett 245(1): 93–98.
Lenzko H, Moog U, Henning K, Lederbach R, Diller R, Menge C, Sachse K, Sprague LD (2011): High frequency of chlamydial co-infections in clinically healthy sheep flocks. BMC Vet Res 7: 29.
Li J, Ma YT, Liang QL, Li RL, Zheng FG, Liu Q, Zhu XQ, Gao WW (2020): Serological evidence of Toxoplasma gondii and Chlamydia infection in alpacas (Vicugna pacos) in Shanxi Province, northern China. Microb Pathog 149: e104399.
Liu B, Zheng DD, Jin Q, Chen LH, Yang J (2019): VFDB 2019: a comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res 47(D1): D687–D692.
Luna EL, Maturrano HL, Rivera GH, Zanabria HV, Rosadio AR (2012): Genotipificación, evaluación toxigénica in vitro y sensibilidad a antibióticos de cepas de Escherichia coli aisladas de casos diarreicos y fatales en alpacas neonatas. Rev Invest Vet Perú 23(3): 280–288.
Malbon AJ, Durrwald R, Kolodziejek J, Nowotny N, Kobera R, Pohle D, Muluneh A, Dervas E, Cebra C, Steffen F, Paternoster G, Gerspach C, Hilbe M (2021): New World camelids are sentinels for the presence of Borna disease virus. Transbound Emerg Dis. doi: 10.1111/tbed.14003.
Maturrano L, Aleman M, Carhuaricra D, Maximiliano J, Siuce J, Luna L, Rosadio R (2018): Draft Genome Sequences of Enterohemorrhagic and Enteropathogenic Escherichia coli Strains Isolated from Alpacas in Peru. Genome Announc 6(1): e01391–01317.
Mercado EC, Rodriguez SM, Elizondo AM, Marcoppido G, Parreno V (2004): Isolation of shiga toxin-producing Escherichia coli from a South American camelid (Lama guanicoe) with diarrhea. J Clin Microbiol 42(10): 4809–4811.
Michelacci V, Tozzoli R, Caprioli A, Martinez R, Scheutz F, Grande L, Sanchez S, Morabito S (2013): A new pathogenicity island carrying an allelic variant of the Subtilase cytotoxin is common among Shiga toxin producing Escherichia coli of human and ovine origin. Clin Microbiol Infec 19(3): E149–E156.
Moennighoff C, Thomas N, Nienhaus F, Hartmann M, Menrath A, Merkel J, Detlefsen H, Kreienbrock L, Hennig-Pauka I (2020): Phenotypic antimicrobial resistance in Escherichia coli strains isolated from swine husbandries in North Western Germany – temporal patterns in samples from laboratory practice from 2006 to 2017. BMC Vet Res 16(1): 37.
Muller D, Greune L, Heusipp G, Karch H, Fruth A, Tschape H, Schmidt MA (2007): Identification of unconventional intestinal pathogenic Escherichia coli isolates expressing intermediate virulence factor profiles by using a novel single-step multiplex PCR. Appl Environ Microbiol 73(10): 3380–3390.
Munster P, Volkel I, von Buchholz A, Czerny CP (2013): Detection of Mycobacterium avium subspecies paratuberculosis by IS900-Based PCR Assays from an Alpaca (Vicugna Pacos) kept in a
German Zoological Garden. J Zoo Wildlife Med 44(1): 176–180.
Neubert S, von Altrock A, Wendt M, Wagener MG (2021): Llama and Alpaca Management in Germany – Results of an Online Survey among Owners on Farm Structure, Health Problems and Self-Reflection. Animals (Basel) 11(1): 102.
Niehaus AJ, Anderson DE (2007): Tooth root abscesses in llamas and alpacas: 123 cases (1994–2005). J Am Vet Med Assocc 231(2): 284–289.
Pantchev A, Sting R, Bauerfeind R, Tyczka J, Sachse K (2010): Detection of all Chlamydophila and Chlamydia spp. of veterinary interest using species-specific real-time PCR assays. Comp Immunol Microb 33(6): 473–484.
Persad AK, LeJeune JT (2014): Animal Reservoirs of Shiga Toxin-Producing Escherichia coli. Microbiol Spectr 2(4): EHEC–0027–2014.
Prkno A, Hoffmann D, Goerigk D, Kaiser M, van Maanen ACF, Jeske K, Jenckel M, Pfaff F, Vahlenkamp TW, Beer M, Ulrich RG, Starke A, Pfeffer M (2017): Epidemiological Investigations of Four Cowpox Virus Outbreaks in Alpaca Herds, Germany. Viruses 9(11): 344.
Probst C (2008): Epidemiologie ausgewählter Infektionskrankheiten von Zooungulaten. Journal-Nr. 3170.
Risco-Castillo V, Wheeler JC, Rosadio R, Garcia-Pena FJ, Arnaiz-Seco I, Hoces D, Castillo H, Veliz A, Ortega-Mora LM (2014): Health impact evaluation of alternative management systems in vicuna (Vicugna vicugna mensalis) populations in Peru. Trop Anim Health Pro 46(4): 641–646.
Salgado M, Herthnek D, Bolske G, Leiva S, Kruze J (2009): First Isolation of Mycobacterium avium subsp paratuberculosis from Wild Guanacos (Lama Guanicoe) on Tierra Del Fuego Island. J Wildlife Dis 45(2): 295–301.
Salgado M, Sevilla I, Rios C, Crossley J, Tejeda C, Manning E (2016): Presence of Mycobacterium avium subsp paratuberculosis in Alpacas (Lama Pacos) Inhabiting the Chilean Altiplano. J Zoo Wildlife Med 47(1): 12–16.
Scheutz F (2014): Taxonomy Meets Public Health: The Case of Shiga Toxin-Producing Escherichia coli. Microbiol Spectr 2(3): EHEC-0019-2013.
Schneeberg A, Rupnik M, Neubauer H, Seyboldt C (2012): Prevalence and distribution of Clostridium difficile PCR ribotypes in cats and dogs from animal shelters in Thuringia, Germany. Anaerobe 18(5): 484–488.
Schulz C, Beer M, Hoffmann B (2015): Schmallenberg virus infection in South American camelids: Field and experimental investigations. Vet Microbiol 180(3-4): 171–179.
Schulze V, Grosse R, Furstenau J, Forth LF, Ebinger A, Richter MT, Tappe D, Mertsch T, Klose K, Schlottau K, Hoffmann B, Hoeper D, Mundhenk L, Ulrich RG, Beer M, Mueller KE, Rubbenstroth D (2020): Borna disease outbreak with high mortality in an alpaca herd in a previously unreported endemic area in Germany. Transbound Emerg Dis 67(5): 2093–2107.
Seemann T (2014): Prokka: rapid prokaryotic genome annotation. Bioinformatics 30(14): 2068–2069.
Shallcross LJ, Davies DS (2014): Antibiotic overuse: a key driver of antimicrobial resistance. Br J Gen Pract 64(629): 604–605.
Shimpoh T, Hirata Y, Ihara S, Suzuki N, Kinoshita H, Hayakawa Y, Ota Y, Narita A, Yoshida S, Yamada A, Koike K (2017): Prevalence of pks-positive Escherichia coli in Japanese patients with or without colorectal cancer. Gut Pathog 9: 35.
Silvera C E, Perales C R, Rodríguez B J, López U T, Gavidia C C, Agapito P J, Palacios E C (2012): Presencia de Escherichia coli O157 en crías de alpacas (Vicugna pacos). Rev Invest Vet Perú 23: 98–104.
Siuce J, Maturrano L, Wheeler JC, Rosadio R (2020): Diarrheagenic Escherichia coli isolates from neonatal alpacas mainly display F17 fimbriae adhesion gene. Trop Anim Health Pro 52(6): 3917–3921.
Stanitznig A, Khol JL, Lambacher B, Franz S, Wittek T, Kralik P, Slana I, Vasickova P (2017): Prevalence of Mycobacterium avium subspecies paratuberculosis and hepatitis E in New World camelids in Austria. Vet Rec 181(2): 46.
Staples P (2016): An introduction to diseases of alpacas in Australia. http://www.flockandherd.net.au/other/reader/diseases-of-alpacas.html (20.06.2021).
Tibary A, Fite C, Anouassi A, Sghiri A (2006): Infectious causes of reproductive loss in camelids. Theriogenology 66(3): 633–647.
Twomey DF, Wu G, Nicholson R, Watson EN, Foster AP (2014): Review of laboratory submissions from New World camelids in England and Wales (2000–2011). Vet J 200(1): 51–59.
Valilis E, Ramsey A, Sidiq S, DuPont HL (2018): Non-O157 Shiga toxin-producing Escherichia coli-A poorly appreciated enteric pathogen: Systematic review. Int J Infect Dis 76: 82–87.
Viesselmann LC, Videla R, Schaefer J, Chapman A, Wyrosdick H, Schaefer DMW (2019): Mycoplasma haemolamae and intestinal parasite relationships with erythrocyte variables in clinically healthy alpacas and llamas. J Vet Intern Med 33(5): 2336–2342.
Whitehead CE, Bedenice D (2009): Neurologic Diseases in Llamas and Alpacas. Vet Clin North Am Food Anim Pract 25(2): 385–405.
Wirth T, Falush D, Lan RT, Colles F, Mensa P, Wieler LH, Karch H, Reeves PR, Maiden MCJ, Ochman H, Achtman M (2006): Sex and virulence in Escherichia coli: an evolutionary perspective. Mol Microbiol 60(5): 1136–1151.
Wood DE, Lu J, Langmead B (2019): Improved metagenomic analysis with Kraken 2. Genome Biol 20(1): 257.
Zankari E, Hasman H, Kaas RS, Seyfarth AM, Agerso Y, Lund O, Larsen MV, Aarestrup FM (2013): Genotyping using whole-genome sequencing is a realistic alternative to surveillance based on phenotypic antimicrobial susceptibility testing. J Antimicrob Chemother 68(4): 771–777.
Zarrin M, Riveros JL, Ahmadpour A, de Almeida AM, Konuspayeva G, Vargas-Bello-Perez E, Faye B, Hernandez-Castellano LE (2020): Camelids: new players in the international animal production context. Trop Anim Health Pro 52(3): 903–913.
Zhou Z, Alikhan NF, Mohamed K, Fan Y, Agama Study G, Achtman M (2020): The EnteroBase user‘s guide, with case studies on Salmonella transmissions, Yersinia pestis phylogeny, and Escherichia core genomic diversity. Genome Res 30(1): 138–152.
Zidaric V, Zemljic M, Janezic S, Kocuvan A, Rupnik M (2008): High diversity of Clostridium difficile genotypes isolated from a single poultry farm producing replacement laying hens. Anaerobe 14(6): 325–327.



Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2021-21-Gonzalez.pdf (0.37 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2021-21-Gonzalez-Table1.pdf (0.07 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2021-21-Gonzalez-Table2.pdf (0.06 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2021-21-Gonzalez-Table3.pdf (0.06 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2021-21-Gonzalez-Table4.pdf (0.06 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF Supplementary_Material_1.xlsx (0.01 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF Supplementary_Material_2.docx (0.04 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF Supplementary_Material_3.xlsx (0.02 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF Supplementary_Material_4.xlsx (0.02 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF Supplementary_Material_5.xlsx (0.01 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF Supplementary_Material_6.xlsx (0.02 MB) herunterladen möchten




{kind=link}
{kind=link}
{kind=link}