

Middle East Respiratory Syndrome Coronavirus (MERS-CoV) – Camel virus and zoonotic pathogen
Berliner und Münchener Tierärztliche Wochenschrift 134, 1-16
DOI: 10.2376/1439-0299-2020-30
© Schlütersche Fachmedien GmbH. 2021
Publiziert: 03/2021
Zusammenfassung
Das im September 2012 erstmals isolierte Middle East Respiratory Syndrome Coronavirus (MERS-CoV) ist die Ursache einer schweren und oft auch tödlich verlaufenden Atemwegserkrankung beim Menschen (Tab. 1). Entsprechend seiner Namensgebung sind MERS-CoV-Infektionen in der Regel auf der saudi-arabischen Halbinsel zu finden. Das MERS-CoV gehört zusammen mit den Severe Acute Respiratory Syndrome-related Coronaviren (SARS-CoV und SARS-CoV-2) zu den zoonotischen Coronaviren, die vom Tier auf den Menschen übertragen werden. Dromedar-Kamele (Camelus dromedarius) stellen ein wichtiges MERS-CoV-Tierreservoir dar, die große Virusmengen mit Sekreten und Exkreten ausscheiden. So kommt es auch zur Übertragung auf den Menschen und zur Aufrechterhaltung des MERS-CoV-Infektionsgeschehens. Bis heute gibt es nur ein begrenztes Wissen zur MERS-CoV-Pathogenese. Zugelassene Impfstoffe und Therapeutika gibt es bisher noch nicht. Die bis heute andauernde MERS-CoV-Epidemie auf der saudi-arabischen Halbinsel zusammen mit der aktuellen SARS-CoV-2-Pandemie verdeutlichen, dass die Entwicklung von Strategien zur effizienten Bekämpfung und Prävention von MERS-CoV und anderen zoonotischen Coronaviren von großer Wichtigkeit für das öffentliche Gesundheitswesen ist.
Summary
The Middle East Respiratory Syndrome Coronavirus (MERS-CoV) has been firstly identified in September 2012 as causative agent of severe and lethal respiratory disease in humans (Table 1). According to its name, MERS-CoV infection mostly occur on the Arabian Peninsula. MERS-CoV together with the Severe acute respiratory syndrome-related coronavirus (SARS-CoV and SARS-CoV-2) are zoonotic coronaviruses that are transmitted by different animals. Dromedary camels (Camelus dromedarius) are an important animal reservoir sheeding high amount of MERS-CoV in nasal discharge indicating a continuous zoonotic transmission in the human population. So far, the understanding of MERS-CoV pathogenesis in humans and camels is very limited. There are no vaccines and therapeutics licensed. However, the MERS-CoV epidemic on the Arabian Peninsula is still ongoing. Moreover, with the ongoing SARS-CoV-2 pandemic, there is another health threat posed by a coronavirus. Thus, the development of efficient therapeutic and preventive strategies is an urgent need for the global health.
Historie
Coronaviren sind schon lange als Ursache verschiedener Erkrankungen bei Mensch und Tier bekannt. Die meisten für die Veterinär- und Humanmedizin relevanten Coronaviren (Tab. 2) verursachen respiratorische und/oder enterische Krankheitsbilder (Cui et al. 2019). Dazu gehören auch die zoonotischen Coronaviren, die vom Tier auf den Menschen übertragen werden.
Ein gut bekannter Vertreter dieser zoonotischen Coronaviren ist das SARS-Coronavirus (engl. Severe Acute Respiratory Syndrome Coronavirus = SARS-CoV), das initial von Fledermäusen über verschiedene mögliche Zwischenwirte auf den Menschen übertragen wurde (Drosten et al. 2003). Das SARS-CoV trat erstmals im Jahr 2002 auf und verursachte eine weltweite Pandemie, an der mehr als 8.000 Menschen erkrankten und 774 starben. Sehr ähnlich dem SARS-CoV ist das SARS-CoV-2, das aktuell die Ursache der COVID-19-Erkrankung ist und zu einer weltweiten Pandemie führte (Hotez et al. 2020). Auch hier wird ein zoonotischer Ursprung vermutet (Andersen et al. 2020). Interessanterweise werden bei Fledermäusen immer wieder Coronaviren gefunden, die eine nahe Verwandtschaft zu Coronaviren beim Menschen aufweisen (Drexler et al. 2014).
Im Jahr 2012 trat erstmals das MERS-Coronavirus (engl. Middle East Respiratory Syndrome Coronavirus = MERS-CoV) als Ursache einer schweren und oftmals auch tödlich verlaufenden Atemwegserkrankung beim Menschen auf (Zaki et al. 2012). So wurde im Juni 2012 ein 60 Jahre alter Mann aus Jeddah in Saudi-
Arabien mit schweren respiratorischen Krankheitssymptomen ins Krankenhaus eingeliefert. Trotz schnell eingeleiteter therapeutischer Maßnahmen und intensivmedizinischer Betreuung verschlimmerte sich die respiratorische Insuffizienz. Darüber hinaus entwickelte der Mann eine begleitende Niereninsuffizienz und verstarb eine Woche später. Aus dem Sputum des Mannes wurde ein neues Coronavirus isoliert, das große Ähnlichkeit zu dem humanen SARS-CoV von 2002/2003 aufwies. Entsprechend der geografischen Region, in welcher das neue Coronavirus zum ersten Mal isoliert wurde, erhielt es den Namen MERS-CoV (de Groot et al. 2013).
Top Job:
Innerhalb kürzester Zeit nach dieser Erstbeschreibung traten weitere MERS-CoV-Infektionen in Saudi-Arabien auf. So wurde im September bei drei weiteren Patienten mit ähnlicher respiratorischer Symptomatik und Nierenerkrankungen ebenfalls das MERS-CoV als Ursache der Erkrankung diagnostiziert. In einer retrospektiven Studie konnte das MERS-CoV außerdem bei einem Ausbruch von schwerer Atemwegssymptomatik im April 2012 in Zarqa in Jordanien identifiziert werden (Al-Abdallat et al. 2014, WHO 2012). Im Gegensatz zum SARS-CoV-Ausbruch im Jahre 2003 dauert das MERS-CoV-Infektionsgeschehen bis heute an. Insgesamt konnte das Virus bisher bei mehr als 2.562 Menschen nachgewiesen werden. 881 Menschen sind infolge einer MERS-CoV-Infektion verstorben (WHO 2019).
Interessanterweise sind bis heute alle Fälle der MERS-CoV-Krankheit epidemiologisch mit der saudi-arabischen Halbinsel in Verbindung zu bringen. So wurden alle größeren MERS-CoV-Ausbrüche im Zusammenhang mit Krankenhaus-assoziierten Clusterausbrüchen in Saudi-Arabien beobachtet (Assiri et al. 2013). Das Hauptcharakteristikum entsprechender Krankenhaus-assoziierter Ausbrüche sind neben fehlenden Hygienemaßnahmen vor allem das Vorhandensein von Menschen mit geschwächtem Immunsystem und Koinfektionen (Bernard-Stoecklin et al. 2019). Die Assoziation einer schweren klinischen Ausprägung der MERS-CoV-Infektion mit Immunsuppression und anderen Begleitinfektionen zeigte sich bereits bei den ersten MERS-CoV-Fällen im Jahre 2012. Hier gab es bis Ende des Jahres neun bestätigte Fälle, von denen fünf Patienten infolge der Infektion sowie Multiorganversagen verstarben (WHO 2012). Bei Menschen mit einem voll funktionsfähigen Immunsystem und gutem Gesundheitszustand verläuft die Erkrankung dagegen in der Regel asymptomatisch oder mild (Alsahafi und Cheng 2016). Bei Infektionen, die in anderen Ländern auftraten, handelte es sich in der Regel um Reisende, welche die saudi-arabische Halbinsel bereist hatten, so auch beim bis dato größten MERS-CoV-Ausbruch außerhalb von Saudi-Arabien, der sich im Jahre 2015 in Südkorea ereignete (Lee und Kim 2016). Hier hatte sich ein südkoreanischer Geschäftsmann während seines Aufenthaltes in Saudi-Arabien mit dem Virus infiziert. Nach seiner Ankunft in Südkorea entwickelte er erste klinische Symptome und wurde im Krankenhaus auf der Intensivstation behandelt. Dort wurde der Mann im Verlauf der weiteren intensivmedizinischen Therapie auch intubiert und, bedingt durch die verzögerte Diagnose, das Virus auf mehrere Krankenpfleger und Ärzte übertragen. Diese übertrugen das MERS-CoV nachfolgend auf weitere Patienten. Insgesamt infizierten sich bei dem Südkorea-Ausbruch 186 Menschen mit MERS-CoV. Durch daraufhin eingeleitete strenge Hygienemaßnahmen einschließlich Patientenisolierung wurde eine weitere Ausbreitung innerhalb von Südkorea und in andere Länder verhindert (Cho et al. 2016, Kim et al. 2017).
Taxonomie und Aufbau des MERS-CoV
Taxonomie
Coronaviren gehören zur Familie der Coronaviridae. Die Coronaviridae gehören zusammen mit der Familie der Arteriviridae zur Ordnung der Nidovirales. Bei den Coronaviren werden aktuell vier Genera (α, β, γ, δ) unterschieden, in welche die jeweiligen Coronaviren entsprechend ihrer phylogenetischen Verwandtschaft eingeteilt werden (Tab. 2). Das MERS-CoV gehört, wie auch die beiden humanen SARS-CoV (SARS-CoV und SARS-CoV-2), zu den β-Coronaviren (de Groot et al. 2013). Das SARS-CoV und SARS-CoV-2 werden in der Gruppe Severe Acute Respiratory Syndrome-related Coronavirus zusammengefasst. Interessanterweise gehören auch die meisten anderen humanmedizinisch relevanten Coronaviren zu dem Genus der β-Coronaviren. Als weitere bekannte Vertreter der humanen Coronaviren sind das Humane Coronavirus NL63 und das Humane Coronavirus 229E zu nennen. Diese sind die Ursache von in der Regel milden Erkältungserkrankungen beim Menschen und gehören zu den α-Coronaviren. Die übrigen Genera umfassen eine Vielzahl von Krankheitserregern, die insbesondere für die Tiermedizin von großer Bedeutung sind.
Manifestationsgewebe im Wirt
Hauptmanifestationsgewebe der Coronaviren sind der Respirationstrakt, der Magen-Darm-Trakt sowie das Nervengewebe inklusive des zentralen Nervensystems (ZNS) (Cui et al. 2019, Masters und Perlman 2013).
Virusaufbau
Bei den Coronaviren handelt es sich um behüllte RNA-Viren mit einem einzelsträngigen Genom von positiver Polarität. Das Genom der Coronaviren ist ca. 30.000 bp groß; damit gehören die Coronaviren zu den größten bisher bekannten RNA-Viren (Masters und Perlman 2013). So besitzt auch das MERS-CoV-Genom eine Größe von 30,1 kb (Abb. 1).
Charakteristisch und namensgebend für die Coronaviren ist das Aussehen des Viruspartikels. In elektronenmikroskopischen Aufnahmen fällt als Erstes die prominente, kronenartige Struktur der Virusoberfläche auf. Diese wird durch das Spikeprotein (S) gebildet. Das keulenförmig erscheinende S-Protein bildet das Hauptstrukturprotein der Coronaviren. Das S-Protein hat eine Größe von 220 kDa; die einzelnen S-Proteine lagern sich jeweils als Trimere zusammen, um die charakteristische Kronenstruktur zu bilden. Die Hauptfunktion des S-Proteins besteht in der Anheftung an den spezifischen Zellrezeptor und der Vermittlung des Viruseintritts in die Zelle. Dementsprechend ist das S-Protein auch das Hauptziel von neutralisierenden Antikörpern. Die Außenseite der Virushülle wird durch das Hüllprotein (engl. envelope = E) gebildet, das sich mit einer Größe von 12 kDa direkt an die Virusmembran anschließt. Die Coronaviren des Genus β-Coronavirus Subgenus Embecovirus besitzen außerdem das Hämagglutininesterase-Protein (HE), das als Transmembranprotein eine wichtige Rolle bei der Anheftung bzw. bei der Abspaltung des durch Knospung aus der infizierten Zelle ausgetretenen neuen Viruspartikels spielt. Die Virusmembran wird von dem Membranprotein (M) gebildet. Das M-Protein weist eine Größe von 35 kDa auf und bildet die Innenseite der Virushülle. Im Innern des Viruspartikels befindet sich das Nukleokapsid, das die virale RNA beinhaltet. Das Nukleokapsid wird hauptsächlich durch das Nukleokapsidprotein (N) gebildet, das eine Größe von 60 kDa besitzt. Das N-Protein steht in direkter Wechselwirkung mit dem M-Protein, um so das Nukleokapsid stabil im Innern des Viruspartikels zu halten. Die im Inneren des Nukleokapsids eingelagerte einzelsträngige RNA ist bei den verschiedenen Vertretern der Coronaviren hochkonserviert (Tab. 3).
Wie die meisten Viren aus der Ordnung der Nidovirales besitzt auch das MERS-CoV eine einzigartige Kodierungsstrategie. Dabei kodieren zwei Drittel des viralen Genoms für zwei große Polyproteine; das restliche Genom wird in einer Reihe von verschiedenen subgenomischen RNAs abgelesen, die damit ein Vielfaches an Kodierungskapazität ermöglichen (Abb. 2).
Das 5‘ Ende des Genoms besitzt eine Cap-Struktur sowie ein einzelnes, großes Open Reading Frame (ORF), das für das Polymerase-Polyprotein kodiert. Daran schließen sich weitere ORFs in Richtung des 3‘ Endes des RNA-Strangs an, die für die weiteren Strukturproteine sowie für akzessorische Proteine kodieren (Masters und Perlman 2013). Das Polyprotein wird durch virale Proteasen in 16 Nichtstrukturproteine (NS) gespalten, die für die Replikation wichtig sind. Diese akzessorischen Proteine stellen eine weitere Form von NS-Proteinen dar, die oftmals eine Bedeutung für die Virulenz aufweisen, wie z. B. durch die spezifische Modulation bzw. Inhibition des Wirtimmunsystems, und werden daher auch Immunevasionsproteine genannt (Channappanavar et al. 2019). Beim MERS-CoV werden folgende akzessorischen Proteine unterschieden: NS3, NS4a, NS4b, NS5, NS8b. In verschiedenen Studien konnte für das MERS-CoV gezeigt werden, dass NS4a und NS4b die Aktivierung der angeborenen Immunantwort und spezifisch den Interferon-Signalweg modulieren (Yang et al. 2013). Allgemein spielen diese akzessorischen Proteine eine wichtige Rolle bei der Virus-Wirt-Interaktion; damit scheinen diese auch wichtig zu sein, um die Infektion empfänglicher Zellen mit dem MERS-CoV zu initiieren (Yang et al. 2015).
Bedeutung der Strukturproteine im Replikationszyklus
Neben dem S-Protein spielen auch die anderen Strukturproteine (M, E, N) eine wichtige Rolle im Verlauf des viralen Vermehrungszyklus (Tab. 3). So vermittelt das M-Protein nach direkter Interaktion mit dem N-Protein die Partikelbildung und das finale Knospen der Virionen. Auch das E-Protein spielt eine Rolle beim Zusammenbau des viralen Partikels. Nach finaler Fusion kommt es zur Abspaltung und Freisetzung der fertigen Viruspartikel von der infizierten Zelle.
Spezifischer Zellrezeptor
Sehr schnell nach der initialen Entdeckung des MERS-CoV wurde auch der spezifische MERS-CoV-Zellrezeptor identifiziert. Anders als die humanen SARS-CoV, die das Angiotensin Converting Enzyme 2 (ACE2) als Zelleintrittsrezeptor nutzen (Hoffmann et al. 2020, Müller et al. 2012), dient für das MERS-CoV die Dipeptidylpeptidase 4 (DPP4) als spezifischer Rezeptor, um den Eintritt des Virus in die Zelle zu vermitteln (Raj et al. 2013). DPP4 kommt auf der Oberfläche von verschiedenen Zellen vor und gehört als proteolytisches Enzym zu den Exopeptidasen. Beim Menschen kommt DPP4 insbesondere auf Zellen des unteren Respirationstrakts, vor allem in der Lunge, sowie in den Nieren vor. Diese Lokalisation erklärt den spezifischen Tropismus und die Klinik von MERS-CoV beim Menschen (Widagdo et al. 2019). Beim Dromedar-Kamel (Camelus dromedarius), einem wichtigen MERS-CoV-Tierreservoir, kommt DPP4 hauptsächlich auf den Zellen des oberen Respirationstraktes vor. Im unteren Respirationstrakt der Kamele konnten keine DPP4 nachgewiesen werden (Widagdo et al. 2016). Dies erklärt die beim Kamel nach MERS-CoV-Infektion auftretende milde respiratorische Infektion, die hauptsächlich auf den oberen Nasen-Rachen-Raum begrenzt ist.
Epidemiologie
Erstbeschreibung
Seit der Erstbeschreibung im Jahr 2012 kommt es regelmäßig zu klinisch manifesten MERS-CoV-Infektionen beim Menschen. Vornehmlich ereignen sich diese Ausbrüche in Saudi-Arabien sowie den übrigen Ländern der Arabischen Halbinsel und des Mittleren Ostens, wie z. B. Bahrain, Irak, Iran, Katar, Oman, Vereinigte Arabische Emirate mit Abu Dhabi, Adschman, Dubai, Fudschaira, Ra’s al-Chaima, Schardscha und Umm al-Qaiwain. Darüber hinaus konnten MERS-CoV-Infektionen beim Menschen bisher in 27 weiteren Ländern diagnostiziert werden, wobei alle diese Fälle epidemiologisch mit einem Kontakt auf der Arabischen Halbinsel in Verbindung gebracht werden konnten (Assiri et al. 2013, Cotten et al. 2014). Der genauere Verlauf der Ausbrüche sowie das aktuelle Infektionsgeschehen wurden bereits im Abschnitt „Historie“ dargestellt.
Tierischer Ursprung
Zum genauen Ursprung des MERS-CoV gibt es bisher noch wenige Daten. Wie auch für die anderen humanen SARS-CoV konnte eine Vielzahl MERS-CoV-ähnlicher Virussequenzen bei Fledermäusen identifiziert werden. Daher fokussierte man sich bei der Suche nach potenziellen MERS-CoV-Reservoirtieren zunächst sehr stark auf Fledermausarten, die auf der saudi-arabischen Halbinsel vorkamen. Hier zeigte sich, dass in Fledermaus-Arten der Familien Vespertilionidae und Molossidae vermeintliche Vorläuferviren des MERS-CoV gefunden werden konnten (Drexler et al. 2014). Dabei konnten jedoch bis heute keine vollständigen Genome dieser Vorläuferviren in den Fledermäusen sequenziert werden, sodass noch keine detaillierten oder abschließenden epidemiologischen Zusammenhänge aufgeklärt wurden.
Im Zuge solcher epidemiologischen Nachforschungen zur Aufklärung von MERS-CoV-Infektionen beim Menschen konnten hohe Titer von neutralisierenden Antikörpern gegen MERS-CoV bei Kamelen im Oman und in Katar detektiert werden. Darüber hinaus gelang es sogar, virale RNA und infektiöses Virus aus dem oberen Respirationstrakt nachzuweisen (Azhar et al. 2014). Weitere serologische Studien bei Kamelen im mittleren Osten bestätigen die hohen Seroprävalenz-Raten für MERS-CoV (Azhar et al. 2014, Haagmans et al. 2014, Kandeil et al. 2019, Raj et al. 2014a, Reusken et al. 2013). Neueste Studien zeigen außerdem, dass auch bei Kamelen in Kenia und im Sudan MERS-CoV-Infektionen diagnostiziert wurden (Farag et al. 2019, Zhang et al. 2019). Diese Studien zeigten auch, dass beim Kamel unterschiedliche genetischen Varianten vorkommen. Dies beinhaltet unter anderem auch Viruslinien, die beim Menschen isoliert werden konnten (Drexler et al. 2014).
Diese Daten implizieren, dass das Dromedar-Kamel ein wichtiges Tierreservoir für das MERS-CoV darstellt (Alsahafi und Cheng 2016). In neuen Studien hat sich gezeigt, dass auch Baktrian-Kamele sowie entsprechende Hybride und Neuweltkameliden eine Empfänglichkeit für das MERS-CoV aufweisen (Adney et al. 2019, Lau et al. 2020). Dies spielt auch epidemiologisch eine nicht zu vernachlässigende Rolle, da in vielen europäischen Ländern (inklusive Deutschland) Lamas und Alpakas gehalten werden. Detaillierte Studien, welche die exakte Rolle der Baktrian-Kamele und der Hybriden sowie der Neuweltkameliden in der MERS-CoV-Epidemiologie aufzeigen, stehen noch aus. Die zoonotische Übertragung des MERS-CoV vom Dromedar-Kamel auf den Menschen kann somit als Ausgangspunkt für ein primäres humanes Infektionsgeschehen angesehen werden (Memish et al. 2014). Diese Hypothese wird weiter erhärtet durch die Omnipräsenz der Kamele im Mittleren Osten sowie den dort gepflegten engen Kontakt zum Menschen und einer Korrelation von MERS-CoV-Infektionen beim Menschen mit vorherigem Kamelkontakt. Dies stellt auch eine mögliche Erklärung dar für das bis heute aktiv andauernde Infektionsgeschehen bei Menschen auf der saudi-arabischen Halbinsel. Dabei kann die höchste Viruslast in Nasensekreten von Dromedar-Kamelen nachgewiesen werden. Dementsprechend erfolgt die effizienteste Übertragung vom Dromedar-Kamel auf den Menschen auch über direkten Kontakt mit diesen Ausscheidungen. So konnte bei Menschen, die beruflich mit Kamelen zu tun haben, im Vergleich zu anderen Berufsgruppen eine höhere Seropositivität detektiert werden. Dies umfasst unter anderem Kamelfarmarbeiter, Kamelschlachthausmitarbeiter und Tierärzte. Dagegen konnte bei diesen Berufsgruppen keine höhere klinische Erkrankungsrate gezeigt werden (Aburizaiza et al. 2014, Alshukairi et al. 2018, Khudhair et al. 2019, Sikkema et al. 2017). Weitere zoonotische Infektionsquellen vom Kamel umfassen rohe, nicht-pasteurisierte Kamelmilch und Fleisch (Hemida et al. 2014, 2015, 2017, Reusken et al. 2014).
Verschiedene phylogenetische Stammbaumanalysen lassen vermuten, dass MERS-CoV initial von Fledermäusen auf Kamele übertragen wurde (Corman et al. 2014). Durch die andauernde Zirkulation in der Kamelpopulation kommt es dort zur Aufrechterhaltung der Infektionskette und nachfolgend zum Überspringen auch auf den Menschen und ggf. zur begrenzten Mensch-zu-Mensch-Übertragung. Hierbei zeigen Sequenzanalysen der beim Menschen isolierten MERS-CoV-Isolate keine signifikanten Sequenzunterschiede im Vergleich zu den beim Kamel isolierten Viren (Cotten et al. 2014, Sabir et al. 2016). Dies spricht dafür, dass MERS-CoV sich nicht spezifisch an den Menschen adaptiert hat, wie es für die humanen SARS-CoV gezeigt werden konnte. Hierbei fällt außerdem auf, dass die zoonotische Übertragung von MERS-CoV vom Kamel auf den Menschen effizienter ist als eine direkte Mensch-zu-Mensch-Übertragung (Adney et al. 2016, Müller et al. 2015, Reusken et al. 2016).
Übertragungsweg
Wichtig für die MERS-Epidemiologie ist auch, dass eine Tröpfchen- und Schmierinfektion über den Mund-Nasen-Rachen-Raum die Hauptübertragungsroute darstellt, was eine relativ effiziente und schnelle Verbreitung des Virus begünstigt (van Doremalen et al. 2013). Für die direkte Mensch-zu-Mensch-Übertragung müssen die betroffenen Individuen in sehr engem Kontakt stehen, damit die beim Menschen tief in der Lunge vorhandenen MERS-CoV-Partikel über Atemwegssekrete übertragen werden können. So zeigen verschiedene Studien, dass bei einem Großteil der primären MERS-CoV-Infektionen Kamele das initiale Virusreservoir waren (Alsahafi und Cheng 2016). Jedoch gibt es darüber hinaus immer noch eine Vielzahl von primären Fällen, bei denen der Ursprung der Infektion nicht ermittelt werden konnte und bis heute Rätsel aufgibt.
Prädispositionsfaktoren
Sekundäre Infektionen beim Menschen stehen häufig im Zusammenhang mit medizinischen Einrichtungen (Drosten et al. 2014, Van Kerkhove et al. 2019). So ereignete sich auch der bislang größte zusammenhängende Ausbruch von MERS-CoV im Frühling 2014 in Ryadh in Saudi-Arabien als Folge eines Krankenhaus-assoziierten Ausbruchs mit mehr als 500 bestätigten Infektionen über das ganze Land verteilt (Fagbo et al. 2015). Im Vergleich zu den anderen humanen β-Coronaviren, wie z. B. SARS-CoV und SARS-CoV-2, besitzt das MERS-CoV eine sehr niedrige Übertragungsrate. Dies zeigt sich insbesondere bei Infektionsketten innerhalb eines Haushaltes. Hier beträgt die Übertragungsrate 4 % (Alsahafi und Cheng 2016, Drosten et al. 2014).
Weitere Studien zeigen, dass verschiedene Prädispositionsfaktoren die Empfänglichkeit für MERS-CoV beim Menschen erhöhen (Alfaraj et al. 2019, Kulcsar et al. 2019). Rund zwei Drittel aller Personen mit bestätigter MERS-CoV-Infektion sind männlich, was auf geschlechtsspezifische Unterschiede bzw. Empfänglichkeit für MERS-CoV hinweist. Eine weitere mögliche Erklärung für diese geschlechtsspezifische Verteilung von MERS-CoV-Infektionen stellt die unterschiedliche Exposition im Mittleren Osten dar (Alsahafi und Cheng 2016, Assiri et al. 2013, Van Kerkhove et al. 2019). Darüber hinaus zeigt die epidemiologische Auswertung der MERS-CoV-Ausbrüche, dass ältere Menschen (> 50 Jahre) ein höheres Risiko besitzen, nach einer Infektion mit MERS-CoV schwer zu erkranken. So besitzen Menschen, die älter als 80 Jahre sind, eine Letalität von 90 % und MERS-CoV-Patienten im Alter von 20–45 Jahren eine Letalität von 10 %.
Ein weiterer Faktor, der die Ausbildung eines schweren und oft auch letalen Krankheitsverlaufs begünstigt, ist das Vorhandensein von Vorerkrankungen wie z. B. Diabetes, Nierenerkrankungen, chronische Herzerkrankungen und chronische Lungenerkrankungen wie Asthma oder chronisch-obstruktive Bronchitis (Kulcsar et al. 2019). Hierbei zeigt sich auch wieder, dass ein Großteil der Ausbruchsgeschehen in Saudi-Arabien mit Krankenhausübertragungen in Verbindung gebracht werden konnte, da die Hauptvirus-Ausscheidung beim Menschen dann erfolgt, wenn klinische Symptome vorhanden sind und die Patienten sich in medizinischer Behandlung befinden (Alsahafi und Cheng 2016).
Pathogenese
Trotz der seit dem Jahr 2012 andauernden MERS-CoV-Epidemie ist das Verständnis über die spezifische Pathogenese des Virus sehr begrenzt. Ein Grund hierfür ist der Mangel an bei MERS-CoV-Patienten durchgeführten Obduktionen sowie von intra vitam entnommenen pathologischen Proben zur Diagnostik und zum histologischen Nachweis von pathologischen Veränderungen und dem In-situ-Nachweis des MERS-CoV. Die begrenzten Daten, die bisher zur MERS-CoV-Pathogenese existieren, stammen überwiegend aus In-vitro-Versuchen sowie aus In-vivo-Studien in verschiedenen MERS-CoV-Tiermodellen (Chan et al. 2013, Hocke et al. 2013, Kindler et al. 2013).
Wie oben beschrieben, stellt das Dromedar ein wichtiges Tierreservoir für das MERS-CoV dar. Entsprechend wurden am Dromedar auch eine Vielzahl von Studien zur Identifizierung der MERS-CoV-Pathogenese durchgeführt (Adney et al. 2016). Eine wichtige Erkenntnis, die auch in anderen Tiermodellen sowie in Zellkulturexperimenten bestätigt werden konnte, war die Identifizierung des zellulären Rezeptors DPP4, wie bereits oben beschrieben.
Bedeutung und Verteilung des DPP4-Rezeptors
DPP4 stellt bisher den am besten verstandenen Mechanismus des MERS-CoV-Tropismus und der Pathogenese dar (Müller et al. 2012, Raj et al. 2014b, Widagdo et al. 2019). Die MERS-CoV-Pathogenese beginnt nach der initialen Infektion der Viren, die primär über den Respirationstrakt erfolgt, mit der spezifischen Bindung des MERS-CoV-S-Proteins an den DPP4-Rezeptor. DPP4 kommt auf verschiedensten Zellen des Respirationstraktes vor.
So wird DPP4 beim Menschen überwiegend auf den Typ-I- und Typ-II-Pneumocyten, auf nicht Zilien tragenden bronchialen Epithelzellen, endothelialen Zellen sowie anderen epithelialen Zellen des unteren Respirationstrakts und der Alveolen exprimiert (Meyerholz et al. 2016, Widagdo et al. 2016). Dabei zeigen verschiedene Studien, dass DPP4 beim Menschen weniger häufig auf den Zellen des oberen Respirationstraktes vorkommt (Boonacker und Van Noorden 2003). Diese Beobachtung stellt eine mögliche Erklärung für die weniger effiziente Mensch-zu-Mensch-Übertragung von MERS-CoV dar.
Effizienter erfolgt eine zoonotische MERS-CoV-Infektion vom Kamel auf den Menschen. Im Gegensatz zum Menschen besitzt das Kamel die DPP4-Moleküle hauptsächlich auf den verschiedenen Zellen des oberen Respirationstraktes. Daher scheidet das Kamel große Mengen von MERS-CoV mit allen respiratorischen Sekreten aus und kann so zur Übertragung auf den Menschen führen (Widagdo et al. 2016).
Nach der primären Infektion beim Menschen müssen die MERS-CoV-Partikel tief in die Lunge gelangen, um dann entsprechend des DPP4-Vorkommens diese Zellen zu infizieren. Darüber hinaus kommt DPP4 auch auf Makrophagen sowie auf epithelialen Zellen von verschiedenen Organen vor. Dazu gehören die Nieren, der Darm, die Leber und der Thymus (Boonacker und Van Noorden 2003). Daher kommt es auch in diesen Organen zur produktiven MERS-CoV-Infektion und der Ausbildung einer spezifischen Klinik. Untersuchungen an Lungen von Patienten mit chronischen Lungenerkrankungen zeigten, dass es hier auf alveolaren Epithelzellen sowie auf den alveolaren Makrophagen zu einer verstärkten Expression von DPP4 kam. Dies stellt eine mögliche Erklärung für die in verschiedenen Studien beschriebene erhöhte Empfänglichkeit einer schweren MERS-CoV-Infektion bei Menschen mit respiratorischen Vorerkrankungen dar (Meyerholz et al. 2016).
Bedeutung der körpereigenen Immunantwort
Neben den direkt durch das Virus hervorgerufenen Schäden und Störungen wird auch immer wieder diskutiert, ob die körpereigene Immunantwort, die als Reaktion auf eine MERS-CoV-Infektion im Individuum aktiviert wird, eine wichtige Rolle bei der spezifischen MERS-CoV-Pathogenese spielt (Zielecki et al. 2013). Dabei ist diese MERS-CoV-spezifische Immunantwort auf der einen Seite wichtig, um die Infektion einzugrenzen und zum Halt zu bringen. Auf der anderen Seite zeigen verschiedene Studien im Kontext von SARS-CoV-Infektionen, dass es durch diese spezifischen Immunreaktionen zur Verschlechterung der Infektion kommen kann. Diese Studien zeigen auch, dass die Aktivierung einer verstärkten angeborenen Immunantwort mit einer frühen und starken Expression von Interferon α/γ sowie verschiedenen Chemokinen häufiger zu einem letalen Ausgang der Infektion führte. Im Gegensatz dazu zeigen SARS-CoV-Patienten, welche die Infektion überlebt haben, eine robuste adaptive Immunantwort (Peiris et al. 2003, Wang et al. 2004).
Diese Daten implizieren, dass das Auftreten einer schweren, letalen Infektion trotz effizienter Aktivierung der angeborenen Immunantwort mit einer Inhibition der adaptiven Immunantwort in Verbindung gebracht werden kann. Weitere Studien, auch im Kontext von MERS-CoV-Infektionen, sind wichtig, um zu untersuchen, ob eine Immunpathogenese auch hier eine Rolle spielt (Drosten et al. 2013).
Klinik
Klinische Symptome beim Dromedar-Kamel
Die MERS-CoV-Infektion beim Menschen und beim Kamel induziert unterschiedliche klinische Bilder. So kommt es beim Kamel in der Regel zur asymptomatischen Klinik bzw. zu einer milden respiratorischen Symptomatik. Hierbei erkranken vornehmlich die jungen Kamele ( 12 Monate) in der immunologischen Lücke, d. h. wenn die maternalen Antikörper abgebaut wurden und noch keine eigene Immunantwort ausgebildet worden ist. In dieser Phase sind die jungen Kamele auch empfänglich für eine MERS-CoV-Infektion (Meyer et al. 2016). Die Übertragung erfolgt hierbei über die Sekrete des oberen Respirationstraktes, wo es entsprechend der Verteilung des DPP4-Rezeptors zur massiven Virusreplikation kommt. Kamele, die das Virus ausscheiden, tragen so zur Aufrechterhaltung der Infektionskette bei (Widagdo et al. 2016).
Spezifische Daten aus Infektionsversuchen beim Kamel, welche die natürlichen Bedingungen in der Kamelpopulation nachahmen, gibt es noch nicht. Vorhandene Daten aus experimentellen Infektionsstudien im Kamel mit einer vermutlich signifikant höheren MERS-CoV-Infektionsdosis im Vergleich zur natürlichen Infektionsdosis zeigen, dass die Kamele nur eine milde klinische Symptomatik des oberen Respirationstrakts mit Nasenausfluss entwickeln (Adney et al. 2016, Haagmans et al. 2016).
Entsprechende Daten aus Feldstudien zeigen, dass es bei den jungen Kamelen in den ersten sechs Monaten nach der Geburt zum Absinken der maternalen Antikörpertiter kommt (Meyer et al. 2016, Saqib et al. 2017). Gleichzeitig konnte MERS-CoV-RNA bei diesen Kamelen nachgewiesen werden, was auf aktive MERS-CoV-Vermehrung hinweist. Im Laufe weiterer sechs Monate kommt es wiederum zum Ansteigen der Antikörper und zur Reduktion der Viruslast bei den Kamelen. Dabei zeigte sich im direkten Vergleich zu den Ergebnissen der adulten Tiere, dass neutralisierende Antikörper beim Kamel nicht zur Induktion einer sterilen Immunität, jedoch zur Reduktion der Virustiter führen und damit eine wichtige Rolle bei der Aufrechterhaltung des MERS-CoV-Infektionsgeschehens innerhalb einer Kamelherde spielen (Meyer et al. 2016). Dies bedingt insbesondere, dass sich die jungen Kamele erstmals infizieren, so das Virus in sehr großen Mengen mit allen Sekreten des oberen Respirationstraktes ausscheiden und scheinbar vor allem in dieser Phase auch auf den Menschen übertragen können.
Bei den adulten Tieren bedingt die nicht sterile Immunität, dass trotz fehlender Symptome und vorhandener neutralisierender Antikörper das Virus zeitweise über den oberen Atmungstrakt ausgeschieden wird und somit dann auch wieder auf die jungen Kamele bzw. den Menschen übertragen werden kann (Adney et al. 2016, Reusken et al. 2016). Diese zoonotische Infektionsquelle stellt neben der Infektion durch junge Kamele vermutlich einen weiteren wichtigen Faktor für die MERS-CoV-Epidemiologie dar.
Klinische Symptome beim Menschen
Beim Menschen können verschiedene klinische Ausprägungen der MERS-CoV-Infektionen unterschieden werden. Diese reichen von einer asymptomatischen bis zu einer schweren, letalen Verlaufsform (Drosten et al. 2013). In der Regel kommt es beim Menschen zu einer milden respiratorischen Erkrankung, die einer Grippe ähnelt (Zumla et al. 2014). Die zoonotische Übertragung vom Kamel auf den Menschen sowie die Mensch-zu-Mensch-Übertragung erfolgen hauptsächlich über die respiratorische Route durch virushaltige Sekrete oder Aerosole (Omrani und Shalhoub 2015).
Nach einer mittleren Inkubationszeit von fünf bis sieben Tagen beginnt die milde Verlaufsform der MERS-CoV-Erkrankung mit grippeähnlichen Symptomen wie leichter Temperaturerhöhung, Halsschmerzen, Schüttelfrost, Kopf- und Gliederschmerzen sowie leichten respiratorischen Symptomen wie Schnupfen und Husten (Assiri et al. 2013, Virlogeux et al. 2016). Hierbei verbessert sich der Zustand nach spätestens fünf Tagen und es kommt zur völligen Ausheilung.
Bei einem erhöhten Risiko für schwere Verläufe, wie z. B. durch Vorerkrankung, Immunsuppression oder erhöhtes Alter, kommt es nach initialer Infektion zu schwereren Verläufen der MERS-CoV-Infektion. Solche Krankheitsverläufe beginnen nach der entsprechenden Inkubationszeit mit klinischen Symptomen, die denen der milden Form ähneln. Hierbei entwickelt sich jedoch sehr rasch eine weiter ansteigende Körpertemperatur bis hin zu hohem Fieber (Virlogeux et al. 2016). Im weiteren Verlauf verschlimmert sich die respiratorische Symptomatik zur respiratorischen Insuffizienz mit schwerem Husten, erhöhter Atemfrequenz und Kurzatmigkeit bis hin zu Atemnot.
In vielen Fällen kommt es außerdem zu Dysfunktionen in anderen Organsystemen, wie beispielsweise kardiovaskuläre Schäden oder Niereninsuffizienz. Eine weitere klinische Ausprägung bei schweren Verlaufsformen umfasst gastrointestinale Symptome mit Bauchschmerzen, Erbrechen und Durchfall. Entsprechend kann es bei solchen Patienten bei kumulierender klinischer Symptomatik zum Multiorganversagen und zum Tod kommen (Al-Tawfiq und Memish 2014, Habib et al. 2019). Die schwere Form der respiratorischen Klinik wird induziert durch eine schwere Pneumonie, die sich im weiteren Verlauf zur respiratorischen Insuffizienz mit akuter Atemnot entwickelt (Arabi et al. 2014).
Hervorzuheben ist außerdem, dass eine Vielzahl der MERS-CoV-Infektionen beim Menschen asymptomatisch ohne signifikante klinische Symptome verlaufen, d. h. in der Regel nicht diagnostiziert werden (Saad et al. 2014). Asymptomatische Infektionen werden häufig erst im Nachhinein bei serologischen Studien aufgedeckt. Eine Studie zur Untersuchung der Seropositivität der Bevölkerung in Saudi-Arabien im Jahr 2014 zeigte, dass ungefähr 45.000 Menschen mit dem MERS-CoV infiziert waren (Müller et al. 2015). Diese Daten sind insbesondere auch wichtig für epidemiologische Studien zur Nachverfolgung von primären Infektionsketten, die bis dato nicht auffindbar waren. Hier lassen sich auch wieder neue direkte Kontakte zu Kamelen als MERS-CoV-Reservoirtiere identifizieren, die bei der originären Kontaktperson aufgrund fehlender Vorerkrankungen oder guter Konstitution des Immunsystems nicht zur klinisch manifesten Infektion geführt hatten, die das Virus dann aber dennoch ausschieden und auf entsprechend empfängliche Individuen weiterverbreiteten. Weitere Studien zur Untersuchung der Seropositivität der saudi-arabischen Bevölkerung werden helfen, die epidemiologischen Zusammenhänge besser zu verstehen.
Laborparameter/Bildgebungsverfahren/Pathologie
Charakteristische Laborparameter einer MERS-CoV-Infektion beinhalten eine akute Lymphozytopenie sowie Thrombozytopenie. Entsprechend der Niereninsuffizienz und Leberschäden können erhöhte Werte bei der Kreatinkinase, Laktatdehydrogenase und erhöhte Alaninaminotransferase sowie Aspartataminotransferase nachgewiesen werden (Alfaraj et al. 2019, Assiri et al. 2013). Beim Röntgenthorax sowie beim Thorax-CT können die durch die MERS-CoV-Infektion induzierten Veränderungen sichtbar gemacht werden (Ajlan et al. 2014). Aufgrund fehlender Obduktionen ist die durch die MERS-CoV-Infektion induzierte Lungenpathologie beim Menschen bisher noch wenig charakterisiert. Eine einzeln durchgeführte Sektion in den Vereinigten Arabischen Emiraten hat gezeigt, dass es vor allem zu einer Verdickung des Lungengewebes inklusive einer generalisierten Blutanstauung und Ödem mit sehr ausgeprägten Schäden an den Alveolen kommt. In der histologischen Untersuchung zeigte sich eine deutliche Entzündung vom exsudativen Typ mit Nekrose der epithelialen Zellen, Hyperplasie der Pneumozyten Typ II, alveolärem Ödem und alveolären Fibrineinlagerungen mit Ablösung der bronchiolären Septen und massiver Einwanderung von Entzündungszellen wie Monozyten/Makrophagen, Neutrophilen und Lymphozyten (Ng et al. 2016).
Bekämpfung
Diagnose
Die bei einer MERS-CoV-Infektion im Vordergrund stehende subklinische oder auch schwere respiratorische Erkrankung beeinträchtigt die Diagnose aufgrund einer Vielzahl an möglichen anderen Differenzialdiagnosen. So kann während der akuten klinischen Phase das MERS-CoV in großen Mengen aus respiratorischen Proben isoliert werden. Besonders geeignet sind Lungenspülproben, die tief aus der Lunge entnommen worden sind. Darüber hinaus lässt sich die virale RNA bisweilen auch aus tiefen Nasenrachentupfern isolieren. Anders verhält es sich beim Dromedar-Kamel; hier reichen Nasentupfer aus dem oberen Respirationstrakt aus, um die MERS-CoV-RNA zu isolieren (Widagdo et al. 2016). In beiden Fällen erfolgt die Diagnose über eine quantitative RT-PCR entweder über ein zweistufiges Verfahren zur Detektion von zwei verschiedenen MERS-CoV-Genen oder direkt mittels einer einzelnen PCR. So werden beim zweistufigen Verfahren Sequenzen oberhalb des E-Gens und oberhalb des ORF-1-Gens nachgewiesen (Corman et al. 2012a). Weitere modifizierte PCR-Reaktionen sind mittlerweile verfügbar. So ist es auch in Anbetracht der aktuellen SARS-CoV-2-Pandemie wichtig, keine Kreuzreaktivität mit anderen humanpathogenen Coronavirusspezies zu detektieren.
Eine weitere Möglichkeit der MERS-CoV-Diagnose stellt die serologische Untersuchung dar. Serum sollte dafür idealerweise zwei bis drei Wochen nach der akuten Infektion gewonnen werden. Hierbei kann entweder über ELISA-Reaktionen der Gehalt an MERS-CoV-spezifischen Antikörpern bestimmt werden, aber auch mittels Serumneutralisationstest oder Plaquereduktionstest (Corman et al. 2012b, Meyer et al. 2014). Eine Schwierigkeit bei der serologischen Diagnostik stellt die geringe Halbwertszeit der MERS-CoV-spezifischen Antikörper dar. So haben verschiedene Studien festgestellt, dass die Antikörpertiter nach überstandener Krankheit sehr schnell abfallen, insbesondere dann, wenn es sich um milde Infektionsverläufe gehandelt hat (Alshukairi et al. 2016, Gierer et al. 2013).
Therapie
Eine spezifische Therapie gegen MERS-CoV-Infektionen gibt es noch nicht, sodass bei der Behandlung symptomatische und begleitende Maßnahmen im Vordergrund stehen (Momattin et al. 2019). Einen Überblick über mögliche Therapeutika bietet Tabelle 4.
Mit antiviralen Wirkstoffen, wie Ribavirin, konnte in verschiedenen Studien ein Effekt auf die MERS-CoV-Infektion erzielt werden. In diesen Studien wurde eine kombinierte Gabe von Ribavirin und dem humanen Cytokin Interferon-α2b (IFN-α2b) auf Wirksamkeit getestet (Falzarano et al. 2013). Ribavirin hemmt spezifisch die virale Polymerase, sodass die Virusreplikation inhibiert wird. Dieser Effekt soll weiter verstärkt werden durch IFN-α2b, das als Typ-I-Interferon ebenfalls eine Inhibition der MERS-CoV-Replikation induziert. In experimentellen Studien konnten nach Gabe von Ribavirin und IFN-α2b früh nach der MERS-CoV-Infektion gute Effekte erzielt werden. So zeigten mit MERS-CoV infizierte Affen eine signifikant reduzierte Viruslast in der Lunge sowie weniger Lungenpathologie (Falzarano et al. 2013). In einer ersten klinischen Testung konnte die Überlebensrate schwer kranker Patienten bei früher Gabe bis auf 14 Tage erhöht werden. Jedoch hatte die Kombinationstherapie aus Ribavirin und IFN-α2b keinen Effekt auf die Überlebensrate schwer kranker MERS-CoV-Patienten, wenn die Gabe zu spät nach Einsetzen der klinischen Symptome erfolgte (Omrani et al. 2014). In einer weiteren Fallstudie, in der die Kombinationstherapie bereits sehr früh nach MERS-CoV-Infektion vor Einsetzen schwerer respiratorischer Symptome begonnen wurde, kam es beim Patienten innerhalb von sechs Tagen zur völligen Genesung ohne weitere Spätfolgen (Al-Tawfiq et al. 2014).
In einem zusätzlichen kombinierten Behandlungsansatz wurde der kombinierte Effekt einer Ribavirin/IFN-α2b- und IFN-β1b-Gabe evaluiert, da sich IFN-β1b als sehr wirksam in In-vitro- und In-vivo-Studien auch in der Monotherapie erwiesen hatte (Morgenstern et al. 2005). Nach der Anwendung bei Patienten, die sich bereits in kritisch krankem Zustand befanden, zeigte die Ribavirin/IFN-α2b/IFN-β1b-Therapie keine besseren Ergebnisse als die Kombination mit IFN-α2b (Shalhoub et al. 2015).
Eine weitere Gruppe bereits etablierter antiviraler Wirkstoffe, die auch gegen Coronaviren untersucht werden, sind die Proteaseinhibitoren. Dazu gehören Lopinavir und Ritonavir, die bereits aus der HIV- sowie SARS-Therapie bekannt sind. Die Wirkungsweise der Protease-Inhibitoren beruht auf der Blockade der viralen Proteasen, die, wie oben beschrieben, die Polyproteine spalten, damit der virale Replikationszyklus ablaufen kann. Dies erfolgt durch inhibitorische Bindung der Proteaseinhibitoren an die spezifischen Substratstellen, die dann nicht mehr funktionsfähig sind. Trotz guter Wirksamkeit gegen das SARS-CoV zeigte Lopinavir keine guten Effekte gegen MERS-CoV in vitro, sodass es bisher nur sehr selten klinisch beim Menschen Anwendung fand. Dagegen zeigte sich wiederum bei einer kombinierten Gabe von Lopinavir und Ritonavir im Affenmodell eine deutliche Verbesserung der klinischen Symptomatik (Chan et al. 2015, Kim et al. 2016). Auf Basis dieser präklinischen Daten soll nun auch eine zukünftige klinische Testung beim Menschen durchgeführt werden.
Eine Vielzahl von weiteren potenziellen Wirkstoffen gegen das MERS-CoV konnte in pharmazeutischen Wirkstoffscreenings identifiziert werden (Hilgenfeld 2014). Dazu gehören beispielsweise Chloroquin, Mykophenolsäure, Chlorpromazin und Nitazoxanid (Cheng et al. 2015, de Wilde et al. 2013, Hart et al. 2014). Diese antiviralen Wirkstoffe müssen aber zunächst noch eine detaillierte präklinische Charakterisierung durchlaufen, bevor sie beim Menschen auf Wirksamkeit getestet werden können.
Ein weiterer vielversprechender Ansatz zur Therapie der MERS-CoV-Infektion beim Menschen stellt die hochdosierte Gabe von MERS-CoV-spezifischen Antikörpern dar.
erschiedene Varianten dieser passiven Immuntherapie sind möglich. So zeigte die Gabe von Plasma von rekonvaleszenten Patienten nach Influenzavirus-Infektionen gute Wirkungen in präklinischen Modellen und auch in ersten klinischen Studien. Ähnliche Ansätze im MERS-CoV-Modell stellen einen vielversprechenden Ansatz zur Therapie dar, sind jedoch schwierig umzusetzen, da es nach MERS-CoV-Infektionen, ähnlich den humanen SARS-CoV-Infektionen, nicht zu hohen und anhaltenden Antikörpertitern kommt. In einer experimentellen Studie zeigte sich, dass es in infizierten Mäusen nach passivem Transfer von Serum von Dromedaren mit hohen Titern von MERS-CoV-spezifischen Antikörpern zur deutlichen Virusreduktion kam (Zhao et al. 2015). Eine weitere Möglichkeit der Isolierung von potenten Antikörpern aus Patienten erfolgte mittels immunologischer Methoden. Diese Antikörper wurden dann weiter amplifiziert und aufgereinigt, sodass diese für die passive Immuntherapie eingesetzt werden konnten. Die präklinische Testung dieser Antikörper in MERS-CoV-infizierten Affen zeigte eine gute Wirkung im Hinblick auf die Viruslast in der Lunge (de Wit et al. 2019, Johnson et al. 2016). In einer weiteren Studie erwies sich ein polyklonaler MERS-CoV-spezifischer Antikörper, der aus Rindern gewonnen wurde, als wirksam in verschiedenen präklinischen Modellen (Luke et al. 2016). Aktuell laufen gerade klinische Studien Phase I zur Testung von IgG-Antikörpergabe im Menschen (Zumla et al. 2016). Auch bei der passiven Gabe von Antikörpern ist, wie auch schon bei der Ribavirin/IFN-α2b-Kombinationstherapie, essenziell, dass die Applikation so früh wie möglich nach der Infektion erfolgt. Dies erweist sich, wie auch bei anderen viral bedingten Erkrankungen, z. B. Influenzavirus, als schwierig, da die ersten klinischen Symptome meist erst dann auftreten, wenn die virale Replikation schon in ausreichendem Maße stattgefunden hat. Daher wird die Verfügbarkeit von wirksamen Impfstoffen gegen das MERS-CoV als sehr vielversprechende Strategie zur Vermeidung von Infektionen angesehen. Solche Impfstoffe können helfen, die MERS-CoV-Infektion gänzlich zu verhindern oder aber auch die klinische Ausprägung der Infektion bei gleichzeitiger Reduktion der Virusausscheidung zu mildern, um dadurch die Übertragungsrate zu vermindern und letztlich zur Ausrottung des Virus beizutragen.
Impfstoffe
Es gibt noch keinen für die Anwendung beim Menschen zugelassenen Impfstoff gegen MERS-CoV. Verschiedene Ansätze zur Entwicklung eines MERS-CoV-Impfstoffes sind aktuell in der klinischen Erprobung am Menschen. Dazu gehören inaktivierte Impfstoffe, attenuierte Lebendimpfstoffe, virale Vektorimpfstoffe, DNA-Impfstoffe und Subunit-Impfstoffe (Fehr et al. 2017, Mubarak et al. 2019, Perlman und Vijay 2016).
Traditionell liegt der Fokus bei der Impfstoffentwicklung gegen Coronaviren auf Strukturproteinen und hier ganz besonders auf dem Spikeprotein, da ein Großteil der Antikörperantwort gegen das Spikeprotein gerichtet ist. Darüber hinaus werden mit Impfstoffen, die auf dem S-Protein basieren auch gute T-Zellantworten aktiviert (Buchholz et al. 2004, He und Jiang 2005, Tripp et al. 2005). Diese Strategie wurde daher auch bei den meisten Impfstoffkandidaten gegen das MERS-CoV verfolgt. So gibt es beispielsweise verschiedene Ansätze zur Entwicklung von viralen Vektorimpfstoffen, die mittels molekularbiologischer Methoden die für das S-Protein kodierenden Sequenzen in den entsprechenden Vektor einbauen. Auch bei den Subunit- und DNA-Impfstoffen basieren die meisten Kandidaten auf dem MERS-CoV-S-Protein. In präklinischen Studien zeigte sich in verschiedenen Tiermodellen eine gute Aktivierung von MERS-CoV-spezifischen Immunantworten, die nach Belastungsinfektion mit dem MERS-CoV auch einen Schutz induzierten (Graham et al. 2013, Perlman und Vijay 2016).
Verschiedene dieser MERS-CoV-Impfstoffkandidaten wurden bereits in klinischen Studien in Phase I und Phase II im Menschen getestet. Hierbei zeigte keiner der getesteten Impfstoffe schwere Nebenwirkungen beim Menschen; alle Impfstoff-Kandidaten induzierten darüber hinaus auch gute Immunantworten gegen das MERS-CoV (Baharoon und Memish 2019, Fehr et al. 2017, Perlman und Vijay 2016). In weiteren klinischen Testungen müssen diese vielversprechenden Impfstoffkandidaten aber noch detaillierter auf Immunogenität, Sicherheit und Wirksamkeit getestet werden. Dies beinhaltet insbesondere die Testung auf die Aktivierung einer Antikörper-abhängigen Verstärkung (Antibody Dependent Enhancement = ADE) der MERS-CoV-Infektion in geimpften Individuen im Vergleich zu nicht-geimpften Patienten. Hinweise auf die Existenz von ADE kommen aus Studien zur Charakterisierung von SARS-CoV-Impfstoffkandidaten. Des Weiteren werden Impfstoffkandidaten gegen Coronaviren historisch bedingt immer mit der Möglichkeit von ADE assoziiert (Hotez et al. 2020, Iwasaki und Yang 2020, Roper und Rehm 2009). Dies basiert auf Daten aus einem Katzenmodell zur Testung eines Impfstoffes, der die Entwicklung von Feliner Infektiöser Peritonitis (FIP) bei der Katze verhindern sollte. Hier kam es infolge der spezifischen Antikörper gegen das Feline Coronavirus bei geimpften Katzen nach Belastungsinfektion zur Entwicklung einer akut schweren klinischen Symptomatik im Vergleich zu den Kontrollgruppen (Scott 1999). Daten zur Existenz von ADE bei MERS-CoV gibt es bisher nicht.
Im Kontext der MERS-CoV-Epidemiologie wird deutlich, dass der Großteil der Bevölkerung mit gutem Gesundheitsstatus aufgrund der milden Infektionsverläufe nicht zur primären Zielpopulation der Impfung gehört. Zielgruppen sind vor allem ältere Menschen und Individuen mit Immunsuppression und Vorerkrankungen, da diese häufig mit schweren klinischen Verläufen erkranken. Dabei stellt sich die Frage, ob man einen möglichen MERS-CoV-Impfstoffkandidaten in einer solchen Risikopopulation überhaupt anwenden könnte, da hier auch die Wahrscheinlichkeit von Impfkomplikationen zunimmt (Simonsen et al. 2007). Weitere wichtige Zielpopulationen für die MERS-CoV-Impfung, die eine wichtige Rolle in der Aufrechterhaltung der Infektionskette spielen, sind Krankenhausmitarbeiter, Kamelfarmarbeiter, Tierärzte und Schlachthausarbeiter. In diesem Zusammenhang stellt auch eine Impfung von Kamelen eine vielversprechende Strategie zur längerfristigen Eradikation des MERS-CoV dar. Dadurch kann nicht nur eine Weiterverbreitung von Kamel zu Kamel, sondern auch eine zoonotische Übertragung auf den Menschen verhindert werden.
Ausblick
Das plötzliche und unerwartete Auftreten des MERS-CoV im Jahr 2012 hat nach dem SARS-CoV 2002/2003 erneut anschaulich verdeutlicht, dass Coronaviren und andere neu oder wiederauftretende Viren eine dauerhafte Bedrohung für den Menschen darstellen. Dies hat sich aktuell mit der seit Dezember 2019 andauernden COVID-19-Pandemie bestätigt. Daher ist es wichtig, auch die MERS-CoV-Epidemie weiter sorgfältig zu beobachten und zu verfolgen sowie die zirkulierenden MERS-CoV auf potenziell auftretende Mutationen zu charakterisieren. Auch die Forschung zum besseren Verständnis pathogenetischer Mechanismen einer MERS-CoV-Infektion ist wichtig, um bessere und effizientere Therapie-Strategien gegen β-Coronaviren zu entwickeln. Hier sind noch viele Fragen und Punkte offen, die auch zum besseren Verständnis der humanen SARS-CoV beitragen können.
Die auf Basis dieser Forschungsfragen etablierten Methoden und Systeme helfen auch im Falle neu auftretender Erreger für eine schnellstmögliche Identifizierung und Charakterisierung. Dies zeigte sich eindrucksvoll mit dem plötzlichen Auftreten von SARS-CoV-2 im Dezember 2019. Trotz der kurzen Zeit und des noch geringen spezifischen Verständnisses gibt es schon viele Daten zu epidemiologischen Fragestellungen sowie erste therapeutische und prophylaktische Ansätze. Viele dieser Ansätze basieren auf Strategien, die erst kürzlich für das MERS-CoV entwickelt wurden und oft auch schon in ersten Studien auf Sicherheit, Effizienz und Wirksamkeit getestet wurden. Dies hat die Entwicklung der SARS-CoV-2-Strategien signifikant beschleunigt, sodass zu hoffen bleibt, dass auch die immer noch laufenden MERS-CoV-Forschungsprojekte weiter bei der Bekämpfung der COVID-19-Pandemie unterstützen.
Ethische Anerkennung
Die Autoren versichern, während des Entstehens der vorliegenden Arbeit die allgemeingültigen Regeln Guter Wissenschaftlicher Praxis befolgt zu haben.
Conflict of interest
Die Autoren versichern, dass keine geschützten, beruflichen oder anderweitigen persönlichen Interessen an einem Produkt oder einer Firma bestehen, welche die in dieser Veröffentlichung genannten Inhalte oder Meinungen beeinflussen können.
Finanzierung
Trifft nicht zu.
Autorenbeitrag
Asisa Volz und Christian Meyer zu Natrup haben das Manuskript gemeinsam erstellt und redigiert.
Korrespondenzadresse
Prof. Dr. Asisa Volz
Institut für Virologie & Research Center
for Emerging Infections and Zoonoses (RIZ)
der Tierärztlichen Hochschule Hannover
Bünteweg 17
30559 Hannover
asisa.volz@tiho-hannover.de
Literatur
Aburizaiza AS, Mattes FM, Azhar EI, Hassan AM, Memish ZA, Muth D, Meyer B, Lattwein E, Müller MA, Drosten C (2014): Investigation of Anti-Middle East Respiratory Syndrome Antibodies in Blood Donors and Slaughterhouse Workers in Jeddah and Makkah, Saudi Arabia, Fall 2012. J Infect Dis 209(2): 243–246. doi: 10.1093/infdis/jit589. Epub 2013 Nov 11.
Adney DR, Bielefeldt-Ohmann H, Hartwig AE, Bowen RA (2016): Infection, Replication, and Transmission of Middle East Respiratory Syndrome Coronavirus in Alpacas. Emerg Infect Dis 22(6): 1031–1037. doi: 10.3201/2206.160192. Epub 2016 Jun 15.
Adney DR, Letko M, Ragan IK, Scott D, Van Doremalen N, Bowen RA, Munster VJ (2019): Bactrian camels shed large quantities of Middle East respiratory syndrome coronavirus (MERS-CoV) after experimental infection. Emerg Microbes Infect 8(1): 717–723. doi: 10.1080/22221751.2019.1618687.
Ajlan AM, Ahyad RA, Jamjoom LG, Alharthy A, Madani TA (2014): Middle East respiratory syndrome coronavirus (MERS-CoV) infection: chest CT findings. Am J Roentgenol 203(4): 782–787. doi: 10.2214/AJR.14.13021.
Al-Abdallat MM, Payne DC, Alqasrawi S, Rha B, Tohme RA, Abedi GR, Al Nsour M, Iblan I, Jarour N, Farag NH, Haddadin A, Al-Sanouri T, Tamin A, Harcourt JL, Kuhar DT, Swerdlow DL, Erdman DD, Pallansch MA, Haynes LM, Gerber SI (2014): Hospital-associated outbreak of Middle East respiratory syndrome coronavirus: a serologic, epidemiologic, and clinical description. Clin Infect Dis 59(9): 1225–1233. doi: 10.1093/cid/ciu359. Epub 2014 May 14.
Al-Tawfiq JA, Memish ZA (2014): Middle East respiratory syndrome coronavirus: epidemiology and disease control measures. Infect Drug Resist 7: 281–287. doi: 10.2147/IDR.S51283. eCollection 2014.
Al-Tawfiq JA, Momattin H, Dib J, Memish ZA (2014): Ribavirin and interferon therapy in patients infected with the Middle East respiratory syndrome coronavirus: an observational study. Int J Infect Dis 20: 42–46. doi: 10.1016/j.ijid.2013.12.003.
Alfaraj SH, Al-Tawfiq JA, Assiri AY, Alzahrani NA, Alanazi AA, Memish ZA (2019): Clinical predictors of mortality of Middle East Respiratory Syndrome Coronavirus (MERS-CoV) infection: A cohort study. Travel Med Infect Dis 29: 48–50. doi: 10.1016/j.tmaid.2019.03.004. Epub 2019 Mar 11.
Alsahafi AJ, Cheng AC (2016): The epidemiology of Middle East respiratory syndrome coronavirus in the Kingdom of Saudi Arabia, 2012–2015. Int J Infect Dis 45: 1–4. doi: 10.1016/j.ijid.2016.02.004. Epub 2016 Feb 10.
Alshukairi A, Khalid I, Ahmed W, Dada A, Bayumi D, Malic L, Althawadi S, Ignacio K, Alsalmi H, Al-Abdely H, Wali G, Qushmaq I, Alraddadi B, Perlman S (2016): Antibody Response and Disease Severity in Healthcare Worker MERS Survivors. Emerg Infect Dis 22(6): 1113–1115. doi: 10.3201/eid2206.160010.
Alshukairi AN, Zheng J, Zhao J, Nehdi A, Baharoon SA, Layqah L, Bokhari A, Al Johani SM, Samman N, Boudjelal M, Ten Eyck P, Al-Mozaini MA, Zhao J, Perlman S, Alagaili AN (2018): High Prevalence of MERS-CoV Infection in Camel Workers in Saudi Arabia. mBio 9(5): e01985-18. doi: 10.1128/mBio.01985-18.
Andersen KG, Rambaut A, Lipkin WI, Holmes C, Garry R (2020): The proximal origin of SARS-CoV-2. Nat Med 26: 450–452. https://doi.org/10.1038/s41591-020-0820-9.
Arabi YM, Arifi AA, Balkhy HH, Najm H, Aldawood AS, Ghabashi A, Hawa H, Alothman A, Khaldi A, Al Raiy B (2014): Clinical course and outcomes of critically ill patients with Middle East respiratory syndrome coronavirus infection. Ann Intern Med 160(6): 389–397. doi: 10.7326/M13-2486.
Assiri A, Mcgeer A, Perl TM, Price CS, Al Rabeeah AA, Cummings DaT, Alabdullatif ZN, Assad M, Almulhim A, Makhdoom H, Madani H, Alhakeem R, Al-Tawfiq JA, Cotten M, Watson SJ, Kellam P, Zumla AI, Memish ZA (2013): Hospital Outbreak of Middle East Respiratory Syndrome Coronavirus. N Engl J Med 369(5): 407–416. doi: 10.1056/NEJMoa1306742. Epub 2013 Jun 19.
Azhar EI, El-Kafrawy SA, Farraj SA, Hassan AM, Al-Saeed MS, Hashem AM, Madani TA (2014): Evidence for Camel-to-Human Transmission of MERS Coronavirus. N Engl J Med 370(26): 2499–2505. doi: 10.1056/NEJMoa1401505. Epub 2014 Jun 4.
Baharoon S, Memish ZA (2019): MERS-CoV as an emerging respiratory illness: A review of prevention methods. Travel Med Infect Dis 32: 101520. doi: 10.1016/j.tmaid.2019.101520.
Bernard-Stoecklin S, Nikolay B, Assiri A, Bin Saeed AA, Ben Embarek PK, El Bushra H, Ki M, Malik MR, Fontanet A, Cauchemez S, Van Kerkhove MD (2019): Comparative Analysis of Eleven Healthcare-Associated Outbreaks of Middle East Respiratory Syndrome Coronavirus (Mers-Cov) from 2015 to 2017. Sci Rep 9(1): 7385. doi: 10.1038/s41598-019-43586-9.
Boonacker E, Van Noorden CJ (2003): The multifunctional or moonlighting protein CD26/DPPIV. Eur J Cell Biol 82(2): 53–73. doi: 10.1078/0171-9335-00302.
Buchholz UJ, Bukreyev A, Yang L, Lamirande EW, Murphy BR, Subbarao K, Collins PL (2004): Contributions of the structural proteins of severe acute respiratory syndrome coronavirus to protective immunity. Proc Natl Acad Sci USA 101(26): 9804–9809. doi: 10.1073/pnas.0403492101. Epub 2004 Jun 21.
Chan JFW, Chan K-H, Kao RYT, To KKW, Zheng B-J, Li CPY, Li PTW, Dai J, Mok FKY, Chen H, Hayden FG, Yuen K-Y (2013): Broad-spectrum antivirals for the emerging Middle East respiratory syndrome coronavirus. J Infect 67(6): 606–616. doi: 10.1016/j.jinf.2013.09.029. Epub 2013 Oct 3.
Chan JFW, Yao Y, Yeung M-L, Deng W, Bao L, Jia L, Li F, Xiao C, Gao H, Yu P, Cai J-P, Chu H, Zhou J, Chen H, Qin C, Yuen K-Y (2015): Treatment With Lopinavir/Ritonavir or Interferon-β1b Improves Outcome of MERS-CoV Infection in a Nonhuman Primate Model of Common Marmoset. J Infect Dis 212(12): 1904–1913. doi: 10.1093/infdis/jiv392. Epub 2015 Jul 21.
Channappanavar R, Fehr AR, Zheng J, Wohlford-Lenane C, Abrahante JE, Mack M, Sompallae R, Mccray PB Jr., Meyerholz DK, Perlman S (2019): IFN-I response timing relative to virus replication determines MERS coronavirus infection outcomes. J Clin Invest 129(9): 3625–3639. doi: 10.1172/JCI126363.
Cheng K-W, Cheng S-C, Chen W-Y, Lin M-H, Chuang S-J, Cheng IH, Sun C-Y, Chou C-Y (2015): Thiopurine analogs and mycophenolic acid synergistically inhibit the papain-like protease of Middle East respiratory syndrome coronavirus. Antiviral Res 115: 9–16. doi: 10.1016/j.antiviral.2014.12.011. Epub 2014 Dec 24.
Cho SY, Kang J-M, Ha YE, Park GE, Lee JY, Ko J-H, Lee JY, Kim JM, Kang C-I, Jo IJ, Ryu JG, Choi JR, Kim S, Huh HJ, Ki C-S, Kang E-S, Peck KR, Dhong H-J, Song J-H, Chung DR, Kim Y-J (2016): MERS-CoV outbreak following a single patient exposure in an emergency room in South Korea: an epidemiological outbreak study. Lancet 388(10048): 994–1001. doi: 10.1016/S0140-6736(16)30623-7. Epub 2016 Jul 9.
Corman VM, Eckerle I, Bleicker T, Zaki A, Landt O, Eschbach-Bludau M, Van Boheemen S, Gopal R, Ballhause M, Bestebroer TM, Muth D, Müller MA, Drexler JF, Zambon M, Osterhaus AD, Fouchier RM, Drosten C (2012a): Detection of a novel human coronavirus by real-time reverse-transcription polymerase chain reaction. Euro Surveill 17(39): 20285. doi: 10.2807/ese.17.39.20285-en.
Corman VM, Müller MA, Costabel U, Timm J, Binger T, Meyer B, Kreher P, Lattwein E, Eschbach-Bludau M, Nitsche A, Bleicker T, Landt O, Schweiger B, Drexler JF, Osterhaus AD, Haagmans BL, Dittmer U, Bonin F, Wolff T, Drosten C (2012b): Assays for laboratory confirmation of novel human coronavirus (hCoV-EMC) infections. Euro Surveill 17(49): 20334. doi: 10.2807/ese.17.49.20334-en.
Corman VM, Ithete NL, Richards LR, Schoeman MC, Preiser W, Drosten C, Drexler JF (2014): Rooting the Phylogenetic Tree of Middle East Respiratory Syndrome Coronavirus by Characterization of a Conspecific Virus from an African Bat. J Virol 88(19): 11297–11303. doi: 10.1128/JVI.01498-14. Epub 2014 Jul 16.
Cotten M, Watson SJ, Zumla AI, Makhdoom HQ, Palser AL, Ong SH, Al Rabeeah AA, Alhakeem RF, Assiri A, Al-Tawfiq JA, Albarrak A, Barry M, Shibl A, Alrabiah FA, Hajjar S, Balkhy HH, Flemban H, Rambaut A, Kellam P, Memish ZA (2014): Spread, Circulation, and Evolution of the Middle East Respiratory Syndrome Coronavirus. mBio 5(1): e01062-13. doi: 10.1128/mBio.01062-13.
Cui J, Li F, Shi Z-L (2019): Origin and evolution of pathogenic coronaviruses. Nat Rev Microbiol 17(3): 181–192. doi: 10.1038/s41579-018-0118-9.
De Groot RJ, Baker SC, Baric RS, Brown CS, Drosten C, Enjuanes L, Fouchier RaM, Galiano M, Gorbalenya AE, Memish ZA, Perlman S, Poon LLM, Snijder EJ, Stephens GM, Woo PCY, Zaki AM, Zambon M, Ziebuhr J (2013): Commentary: Middle East Respiratory Syndrome Coronavirus (MERS-CoV): Announcement of the Coronavirus Study Group. J Virol 87(14): 7790–7792. doi: 10.1128/JVI.01244-13. Epub 2013 May 15.
De Wilde AH, Raj VS, Oudshoorn D, Bestebroer TM, Van Nieuwkoop S, Limpens RWaL, Posthuma CC, Van Der Meer Y, Bárcena M, Haagmans BL, Snijder EJ, Van Den Hoogen BG (2013): MERS-coronavirus replication induces severe in vitro cytopathology and is strongly inhibited by cyclosporin A or interferon-α treatment. J Gen Virol 94(Pt 8): 1749–1760. doi: 10.1099/vir.0.052910-0. Epub 2013 Apr 25.
De Wit E, Feldmann F, Horne E, Okumura A, Cameroni E, Haddock E, Saturday G, Scott D, Gopal R, Zambon M, Corti D, Feldmann H (2019): Prophylactic efficacy of a human monoclonal antibody against MERS-CoV in the common marmoset. Antiviral Res 163: 70–74. doi: 10.1016/j.antiviral.2019.01.016. Epub 2019 Jan 24.
De Wit E, van Doremalen N, Falzarono D, Munster VJ (2016): SARS and MERS: recent insights into emerging coronaviruses. Nat Rev Microbiol 14(8): 523–534. doi: 10.1038/nrmicro.2016.81. Epub 2016 Jun 27.
Drexler JF, Corman VM, Drosten C (2014): Ecology, evolution and classification of bat coronaviruses in the aftermath of SARS. Antiviral Res 101: 45–56. doi: 10.1016/j.antiviral.2013.10.013. Epub 2013 Oct 31.
Drosten C, Günther S, Preiser W, Van Der Werf S, Brodt H-R, Becker S, Rabenau H, Panning M, Kolesnikova L, Fouchier RaM, Berger A, Burguière A-M, Cinatl J, Eickmann M, Escriou N, Grywna K, Kramme S, Manuguerra J-C, Müller S, Rickerts V, Stürmer M, Vieth S, Klenk H-D, Osterhaus ADME, Schmitz H, Doerr HW (2003): Identification of a Novel Coronavirus in Patients with Severe Acute Respiratory Syndrome. N Engl J Med 348(20): 1967–1976. doi: 10.1056/NEJMoa030747. Epub 2003 Apr 10.
Drosten C, Seilmaier M, Corman VM, Hartmann W, Scheible G, Sack S, Guggemos W, Kallies R, Muth D, Junglen S, Müller MA, Haas W, Guberina H, Röhnisch T, Schmid-Wendtner M, Aldabbagh S, Dittmer U, Gold H, Graf P, Bonin F, Rambaut A, Wendtner C-M (2013): Clinical features and virological analysis of a case of Middle East respiratory syndrome coronavirus infection. Lancet Infect Dis 13(9): 745–751. doi: 10.1016/S1473-3099(13)70154-3. Epub 2013 Jun 17.
Drosten C, Meyer B, Müller MA, Corman VM, Al-Masri M, Hossain R, Madani H, Sieberg A, Bosch BJ, Lattwein E, Alhakeem RF, Assiri AM, Hajomar W, Albarrak AM, Al-Tawfiq JA, Zumla AI, Memish ZA (2014): Transmission of MERS-Coronavirus in Household Contacts. N Engl J Med 371(9): 828–835. doi: 10.1056/NEJMoa1405858.
Fagbo S, Skakni L, Chu DKW, Garbati M, Joseph M, Hakawi A (2015): Molecular Epidemiology of Hospital Outbreak of Middle East Respiratory Syndrome, Riyadh, Saudi Arabia, 2014. Emerg Infect Dis 21(11): 1981–1988. doi: 10.3201/eid2111.150944.
Falzarano D, De Wit E, Rasmussen AL, Feldmann F, Okumura A, Scott DP, Brining D, Bushmaker T, Martellaro C, Baseler L, Benecke AG, Katze MG, Munster VJ, Feldmann H (2013): Treatment with interferon-α2b and ribavirin improves outcome in MERS-CoV-infected rhesus macaques. Nat Med 19(10): 1313–1317. doi: 10.1038/nm.3362. Epub 2013 Sep 8.
Farag E, Sikkema RS, Mohamedani AA, De Bruin E, Munnink BBO, Chandler F, Kohl R, Van Der Linden A, Okba NMA, Haagmans BL, Van Den Brand JMA, Elhaj AM, Abakar AD, Nour BYM, Mohamed AM, Alwaseela BE, Ahmed H, Alhajri MM, Koopmans M, Reusken C, Elrahman SHA (2019): MERS-CoV in Camels but Not Camel Handlers, Sudan, 2015 and 2017. Emerg Infect Dis 25(12): 2333–2335. doi: 10.3201/eid2512.190882.
Fehr AR, Channappanavar R, Perlman S (2017): Middle East Respiratory Syndrome: Emergence of a Pathogenic Human Coronavirus. Annu Rev Med 68: 387–399. doi: 10.1146/annurev-med-051215-031152. Epub 2016 Aug 26.
Gierer S, Hofmann-Winkler H, Albuali W, Bertram S, Al-Rubaish A, Yousef A, Al-Nafaie A, Al-Ali A, Obeid O, Alkharsah K, Pöhlmann S (2013): Lack of MERS Coronavirus Neutralizing Antibodies in Humans, Eastern Province, Saudi Arabia. Emerg Infect Dis 19(12): 2034–2036. doi: 10.3201/eid1912.130701.
Graham RL, Donaldson EF, Baric RS (2013): A decade after SARS: strategies for controlling emerging coronaviruses. Nat Rev Microbiol 11(12): 836–848. doi: 10.1038/nrmicro3143. Epub 2013 Nov 11.
Haagmans BL, Al Dhahiry SH, Reusken CB, Raj VS, Galiano M, Myers R, Godeke GJ, Jonges M, Farag E, Diab A, Ghobashy H, Alhajri F, Al-Thani M, Al-Marri SA, Al Romaihi HE, Al Khal A, Bermingham A, Osterhaus AD, Alhajri MM, Koopmans MP (2014): Middle East respiratory syndrome coronavirus in dromedary camels: an outbreak investigation. Lancet Infect Dis 14(2): 140–145. doi: 10.1016/S1473-3099(13)70690-X. Epub 2013 Dec 17.
Haagmans BL, Van Den Brand JM, Raj VS, Volz A, Wohlsein P, Smits SL, Schipper D, Bestebroer TM, Okba N, Fux R, Bensaid A, Solanes Foz D, Kuiken T, Baumgärtner W, Segalés J, Sutter G, Osterhaus AD (2016): An orthopoxvirus-based vaccine reduces virus excretion after MERS-CoV infection in dromedary camels. Science 351(6268): 77–81. doi: 10.1126/science.aad1283. Epub 2015 Dec 17.
Haake C, Cook S, Pusterla N, Murphy B (2020): Coronavirus Infections in Companion Animals: Virology, Epidemiology, Clinical and Pathologic Features. Viruses 12(9): 1023. doi: 10.3390/v12091023.
Habib AMG, Ali MaE, Zouaoui BR, Taha MaH, Mohammed BS, Saquib N (2019): Clinical outcomes among hospital patients with Middle East respiratory syndrome coronavirus (MERS-CoV) infection. BMC Infect Dis 19(1): 870. doi: 10.1186/s12879-019-4555-5.
Hart BJ, Dyall J, Postnikova E, Zhou H, Kindrachuk J, Johnson RF, Olinger GG, Frieman MB, Holbrook MR, Jahrling PB, Hensley L (2014): Interferon-β and mycophenolic acid are potent inhibitors of Middle East respiratory syndrome coronavirus in cell-based assays. J Gen Virol 95(Pt 3): 571–577. doi: 10.1099/vir.0.061911-0. Epub 2013 Dec 9.
He Y, Jiang S (2005): Vaccine design for severe acute respiratory syndrome coronavirus. Viral Immunol 18(2): 327–332. doi: 10.1089/vim.2005.18.327.
Hemida M, Chu DKW, Poon LLM, Perera RaPM, Alhammadi MA, Ng H-Y, Siu LY, Guan Y, Alnaeem A, Peiris M (2014): MERS Coronavirus in Dromedary Camel Herd, Saudi Arabia. Emerg Infect Dis 20(7): 1231–1234. doi: 10.3201/eid2007.140571.
Hemida M, Al-Naeem A, Perera RaPM, Chin AWH, Poon LLM, Peiris M (2015): Lack of Middle East Respiratory Syndrome Coronavirus Transmission from Infected Camels. Emerg Infect Dis 21(4): 699–701. doi: 10.3201/eid2104.141949.
Hemida M, Elmoslemany A, Al-Hizab F, Alnaeem A, Almathen F, Faye B, Chu DKW, Perera RaPM, Peiris M (2017): Dromedary Camels and the Transmission of Middle East Respiratory Syndrome Coronavirus (MERS-CoV). Transbound Emerg Dis 64(2): 344–353. doi: 10.1111/tbed.12401. Epub 2015 Aug 10.
Hilgenfeld R (2014): From SARS to MERS: crystallographic studies on coronaviral proteases enable antiviral drug design. FEBS J 281(18): 4085–4096. doi: 10.1111/febs.12936. Epub 2014 Aug 11.
Hocke AC, Becher A, Knepper J, Peter A, Holland G, Tönnies M, Bauer TT, Schneider P, Neudecker J, Muth D, Wendtner CM, Rückert JC, Drosten C, Gruber AD, Laue M, Suttorp N, Hippenstiel S, Wolff T (2013): Emerging Human Middle East Respiratory Syndrome Coronavirus Causes Widespread Infection and Alveolar Damage in Human Lungs. Am J Respir Crit Care Med 188(7): 882–886. doi: 10.1164/rccm.201305-0954LE.
Hoffmann M, Kleine-Weber H, Schroeder S, Krüger N, Herrler T, Erichsen S, Schiergens TS, Herrler G, Wu N-H, Nitsche A, Müller MA, Drosten C, Pöhlmann S (2020): SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 181(2): 271–280.e8. doi: 10.1016/j.cell.2020.02.052. Epub 2020 Mar 5.
Hotez PJ, Corry DB, Bottazzi ME (2020): COVID-19 vaccine design: the Janus face of immune enhancement. Nat Rev Immunol 20(6): 347–348. doi: 10.1038/s41577-020-0323-4. Epub 2020 Apr 28.
Iwasaki A, Yang Y (2020): The potential danger of suboptimal antibody responses in COVID-19. Nat Rev Immunol 20(6): 339–341. doi: 10.1038/s41577-020-0321-6.
Johnson RF, Bagci U, Keith L, Tang X, Mollura DJ, Zeitlin L, Qin J, Huzella L, Bartos CJ, Bohorova N, Bohorov O, Goodman C, Kim DH, Paulty MH, Velasco J, Whaley KJ, Johnson JC, Pettitt J, Ork BL, Solomon J, Oberlander N, Zhu Q, Sun J, Holbrook MR, Olinger GG, Baric RS, Hensley LE, Jahrling PB, Marasco WA (2016): 3B11-N, a monoclonal antibody against MERS-CoV, reduces lung pathology in rhesus monkeys following intratracheal inoculation of MERS-CoV Jordan-n3/2012. Virology 490: 49–58. doi: 10.1016/j.virol.2016.01.004. Epub 2016 Jan 30.
Kandeil A, Gomaa M, Nageh A, Shehata MM, Kayed AE, Sabir JSM, Abiadh A, Jrijer J, Amr Z, Said MA, Byarugaba DK, Wabwire-Mangen F, Tugume T, Mohamed NS, Attar R, Hassan SM, Linjawi SA, Moatassim Y, Kutkat O, Mahmoud S, Bagato O, Shama NMA, El-Shesheny R, Mostafa A, Perera RA, Chu DK, Hassan N, Elsokary B, Saad A, Sobhy H, El Masry I, Mckenzie PP, Webby RJ, Peiris M, Makonnen YJ, Ali MA, Kayali G (2019): Middle East Respiratory Syndrome Coronavirus (MERS-CoV) in Dromedary Camels in Africa and Middle East. Viruses 11(8): 717. doi: 10.3390/v11080717.
Khudhair A, Killerby M, Al Mulla M, Abou Elkheir K, Ternanni W, Bandar Z, Weber S, Khoury M, Donnelly G, Al Muhairi S, Khalafalla A, Trivedi S, Tamin A, Thornburg N, Watson J, Gerber S, Al Hosani F, Hall A (2019): Risk Factors for MERS-CoV Seropositivity among Animal Market and Slaughterhouse Workers, Abu Dhabi, United Arab Emirates, 2014–2017. Emerg Infect Dis 25(5): 927–935. doi: 10.3201/eid2505.181728.
Kim KH, Tandi TE, Choi JW, Moon JM, Kim MS (2017): Middle East respiratory syndrome coronavirus (MERS-CoV) outbreak in South Korea, 2015: epidemiology, characteristics and public health implications. J Hosp Infect 95(2): 207–213. doi: 10.1016/j.jhin.2016.10.008. Epub 2016 Oct 14.
Kim UJ, Won EJ, Kee SJ, Jung SI, Jang HC (2016): Combination therapy with lopinavir/ritonavir, ribavirin and interferon-α for Middle East respiratory syndrome. Antivir Ther 21(5): 455–459. doi: 10.3851/IMP3002. Epub 2015 Oct 22.
Kindler E, Jónsdóttir HR, Muth D, Hamming OJ, Hartmann R, Rodriguez R, Geffers R, Fouchier RaM, Drosten C, Müller MA, Dijkman R, Thiel V (2013): Efficient Replication of the Novel Human Betacoronavirus EMC on Primary Human Epithelium Highlights Its Zoonotic Potential. mBio 4(1): e00611-12. doi: 10.1128/mBio.00611-12.
Kulcsar KA, Coleman CM, Beck SE, Frieman MB (2019): Comorbid diabetes results in immune dysregulation and enhanced disease severity following MERS-CoV infection. JCI Insight 4(20): e131774. doi: 10.1172/jci.insight.131774.
Lau SKP, Li KSM, Luk HKH, He Z, Teng JLL, Yuen K-Y, Wernery U, Woo PCY (2020): Middle East Respiratory Syndrome Coronavirus Antibodies in Bactrian and Hybrid Camels from Dubai. mSphere 5(1): e00898-19. doi: 10.1128/mSphere.00898-19.
Lee J, Kim WJ (2016): Collaborative Intervention of Middle East Respiratory Syndrome: Rapid Response Team. Infect Chemother 48(2): 71–74. doi: 10.3947/ic.2016.48.2.71. Epub 2016 Jun 30.
Luke T, Wu H, Zhao J, Channappanavar R, Coleman CM, Jiao J-A, Matsushita H, Liu Y, Postnikova EN, Ork BL, Glenn G, Flyer D, Defang G, Raviprakash K, Kochel T, Wang J, Nie W, Smith G, Hensley LE, Olinger GG, Kuhn JH, Holbrook MR, Johnson RF, Perlman S, Sullivan E, Frieman MB (2016): Human polyclonal immunoglobulin G from transchromosomic bovines inhibits MERS-CoV in vivo. Sci Transl Med 8(326): 326ra21. doi: 10.1126/scitranslmed.aaf1061.
Masters P, Perlman S (2013): Coronaviridae. In: Fields B, Knipe DM, Howley PM (eds.), Fields Virology. 6th ed. Lippincott Williams & Wilkins, Philadelphia, PA, 825–858.
Memish Z, Cotten M, Meyer B, Watson S, Alsahafi A, Al Rabeeah A, Corman VM, Sieberg A, Makhdoom H, Assiri A, Al Masri M, Aldabbagh S, Bosch B-J, Beer M, Müller M, Kellam P, Drosten C (2014): Human Infection with MERS Coronavirus after Exposure to Infected Camels, Saudi Arabia, 2013. Emerg Infect Dis 20(6): 1012–1015. doi: 10.3201/eid2006.140402.
Meyer B, Drosten C, Müller MA (2014): Serological assays for emerging coronaviruses: Challenges and pitfalls. Virus Res 194: 175–183. doi: 10.1016/j.virusres.2014.03.018. Epub 2014 Mar 23.
Meyer B, Juhasz J, Barua R, Das Gupta A, Hakimuddin F, Corman VM, Müller MA, Wernery U, Drosten C, Nagy P (2016): Time Course of MERS-CoV Infection and Immunity in Dromedary Camels. Emerg Infect Dis 22(12): 2171–2173. doi: 10.3201/eid2212.160382. Epub 2016 Dec 15.
Meyerholz DK, Lambertz AM, Mccray PB (2016): Dipeptidyl Peptidase 4 Distribution in the Human Respiratory Tract: Implications for the Middle East Respiratory Syndrome. Am J Pathol 186(1): 78–86. doi: 10.1016/j.ajpath.2015.09.014. Epub 2015 Nov 18.
Momattin H, Al-Ali AY, Al-Tawfiq JA (2019): A Systematic Review of therapeutic agents for the treatment of the Middle East Respiratory Syndrome Coronavirus (MERS-CoV). Travel Med Infect Dis 30: 9–18. doi: 10.1016/j.tmaid.2019.06.012. Epub 2019 Jun.
Morgenstern B, Michaelis M, Baer PC, Doerr HW, Cinatl J Jr. (2005): Ribavirin and interferon-beta synergistically inhibit SARS-associated coronavirus replication in animal and human cell lines. Biochem Biophys Res Commun 326(4): 905–908. doi: 10.1016/j.bbrc.2004.11.128.
Mubarak A, Alturaiki W, Hemida MG (2019): Middle East Respiratory Syndrome Coronavirus (MERS-CoV): Infection, Immunological Response, and Vaccine Development. J Immunol Res 2019: 6491738. doi: 10.1155/2019/6491738. eCollection 2019.
Müller MA, Raj VS, Muth D, Meyer B, Kallies S, Smits SL, Wollny R, Bestebroer TM, Specht S, Suliman T, Zimmermann K, Binger T, Eckerle I, Tschapka M, Zaki AM, Osterhaus ADME, Fouchier RaM, Haagmans BL, Drosten C (2012): Human Coronavirus EMC Does Not Require the SARS-Coronavirus Receptor and Maintains Broad Replicative Capability in Mammalian Cell Lines. mBio 3(6): e00515-12. doi: 10.1128/mBio.00515-12.
Müller MA, Meyer B, Corman VM, Al-Masri M, Turkestani A, Ritz D, Sieberg A, Aldabbagh S, Bosch B-J, Lattwein E, Alhakeem RF, Assiri AM, Albarrak AM, Al-Shangiti AM, Al-Tawfiq JA, Wikramaratna P, Alrabeeah AA, Drosten C, Memish ZA (2015): Presence of Middle East respiratory syndrome coronavirus antibodies in Saudi Arabia: a nationwide, cross-sectional, serological study. Lancet Infect Dis 15(5): 559–564. doi: 10.1016/S1473-3099(15)70090-3. Epub 2015 Apr 8.
Ng DL, Al Hosani F, Keating MK, Gerber SI, Jones TL, Metcalfe MG, Tong S, Tao Y, Alami NN, Haynes LM, Mutei MA, Abdel-Wareth L, Uyeki TM, Swerdlow DL, Barakat M, Zaki SR (2016): Clinicopathologic, Immunohistochemical, and Ultrastructural Findings of a Fatal Case of Middle East Respiratory Syndrome Coronavirus Infection in the United Arab Emirates, April 2014. Am J Pathol 186(3): 652–658. doi: 10.1016/j.ajpath.2015.10.024. Epub 2016 Feb 5.
Omrani AS, Shalhoub S (2015): Middle East respiratory syndrome coronavirus (MERS-CoV): what lessons can we learn? J Hosp Infect 91(3): 188–196. doi: 10.1016/j.jhin.2015.08.002. Epub 2015 Aug 22.
Omrani AS, Saad MM, Baig K, Bahloul A, Abdul-Matin M, Alaidaroos AY, Almakhlafi GA, Albarrak MM, Memish ZA, Albarrak AM (2014): Ribavirin and interferon alfa-2a for severe Middle East respiratory syndrome coronavirus infection: a retrospective cohort study. Lancet Infect Dis 14(11): 1090–1095. doi: 10.1016/S1473-3099(14)70920-X. Epub 2014 Sep 29.
Peiris JSM, Chu CM, Cheng VCC, Chan KS, Hung IFN, Poon LLM, Law KI, Tang BSF, Hon TYW, Chan CS, Chan KH, Ng JSC, Zheng BJ, Ng WL, Lai RWM, Guan Y, Yuen KY (2003): Clinical progression and viral load in a community outbreak of coronavirus-associated SARS pneumonia: a prospective study. Lancet 361(9371): 1767–1772. doi: 10.1016/s0140-6736(03)13412-5.
Perlman S, Vijay R (2016): Middle East respiratory syndrome vaccines. Int J Infect Dis 47: 23–28. doi: 10.1016/j.ijid.2016.04.008. Epub 2016 Apr 7.
Raj VS, Mou H, Smits SL, Dekkers DHW, Müller MA, Dijkman R, Muth D, Demmers JaA, Zaki A, Fouchier RaM, Thiel V, Drosten C, Rottier PJM, Osterhaus ADME, Bosch BJ, Haagmans BL (2013): Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature 495(7440): 251–254. doi: 10.1038/nature12005.
Raj VS, Farag EaBA, Reusken CBEM, Lamers M, Pas S, Voermans J, Smits S, Osterhaus ADME, Al-Mawlawi N, Al-Romaihi H, Alhajri M, El-Sayed A, Mohran K, Ghobashy H, Alhajri F, Al-Thani M, Al-Marri S, El-Maghraby M, Koopmans MPG, Haagmans B (2014a): Isolation of MERS Coronavirus from a Dromedary Camel, Qatar, 2014. Emerg Infect Dis 20(8): 1339–1342. doi: 10.3201/eid2008.140663.
Raj VS, Smits SL, Provacia LB, Van Den Brand JMA, Wiersma L, Ouwendijk WJD, Bestebroer TM, Spronken MI, Van Amerongen G, Rottier PJM, Fouchier RaM, Bosch BJ, Osterhaus ADME, Haagmans BL (2014b): Adenosine Deaminase Acts as a Natural Antagonist for Dipeptidyl Peptidase 4-Mediated Entry of the Middle East Respiratory Syndrome Coronavirus. J Virol 88(3): 1834–1838. doi: 10.1128/JVI.02935-13. Epub 2013 Nov 20.
Reusken CB, Haagmans BL, Müller MA, Gutierrez C, Godeke G-J, Meyer B, Muth D, Raj VS, Vries LS-D, Corman VM, Drexler J-F, Smits SL, El Tahir YE, De Sousa R, Van Beek J, Nowotny N, Van Maanen K, Hidalgo-Hermoso E, Bosch B-J, Rottier P, Osterhaus A, Gortázar-Schmidt C, Drosten C, Koopmans MPG (2013): Middle East respiratory syndrome coronavirus neutralising serum antibodies in dromedary camels: a comparative serological study. Lancet Infect Dis 13(10): 859–866. doi: 10.1016/S1473-3099(13)70164-6. Epub 2013 Aug 9.
Reusken CB, Farag EA, Jonges M, Godeke GJ, El-Sayed AM, Pas SD, Raj VS, Mohran KA, Moussa HA, Ghobashy H, Alhajri F, Ibrahim AK, Bosch BJ, Pasha SK, Al-Romaihi HE, Al-Thani M, Al-Marri SA, Alhajri MM, Haagmans BL, Koopmans MP (2014): Middle East respiratory syndrome coronavirus (MERS-CoV) RNA and neutralising antibodies in milk collected according to local customs from dromedary camels, Qatar, April 2014. Euro Surveill 19(23): 20829. doi: 10.2807/1560-7917.es2014.19.23.20829.
Reusken CB, Raj VS, Koopmans MP, Haagmans BL (2016): Cross host transmission in the emergence of MERS coronavirus. Curr Opin Virol 16: 55–62. doi: 10.1016/j.coviro.2016.01.004. Epub 2016 Jan 29.
Roper RL, Rehm KE (2009): SARS vaccines: where are we? Expert Rev Vaccines 8(7): 887–898. doi: 10.1586/erv.09.43.
Saad M, Omrani AS, Baig K, Bahloul A, Elzein F, Matin MA, Selim MA, Al Mutairi M, Al Nakhli D, Al Aidaroos AY, Al Sherbeeni N, Al-Khashan HI, Memish ZA, Albarrak AM (2014): Clinical aspects and outcomes of 70 patients with Middle East respiratory syndrome coronavirus infection: a single-center experience in Saudi Arabia. Int J Infect Dis 29: 301–306. doi: 10.1016/j.ijid.2014.09.003. Epub 2014 Oct 7.
Sabir JSM, Lam TTY, Ahmed MMM, Li L, Shen Y, Abo-Aba SEM, Qureshi MI, Abu-Zeid M, Zhang Y, Khiyami MA, Alharbi NS, Hajrah NH, Sabir MJ, Mutwakil MHZ, Kabli SA, Alsulaimany FaS, Obaid AY, Zhou B, Smith DK, Holmes EC, Zhu H, Guan Y (2016): Co-circulation of three camel coronavirus species and recombination of MERS-CoVs in Saudi Arabia. Science 351(6268): 81–84. doi: 10.1126/science.aac8608. Epub 2015 Dec 17.
Saqib M, Sieberg A, Hussain MH, Mansoor MK, Zohaib A, Lattwein E, Müller MA, Drosten C, Corman VM (2017): Serologic Evidence for MERS-CoV Infection in Dromedary Camels, Punjab, Pakistan, 2012–2015. Emerg Infect Dis 23(3): 550–551. doi: 10.3201/eid2303.161285.
Scott FW (1999): Evaluation of risks and benefits associated with vaccination against coronavirus infections in cats. Adv Vet Med 1999;41:347-58. doi: 10.1016/s0065-3519(99)80026-3.
Shalhoub S, Farahat F, Al-Jiffri A, Simhairi R, Shamma O, Siddiqi N, Mushtaq A (2015): IFN-α2a or IFN-β1a in combination with ribavirin to treat Middle East respiratory syndrome coronavirus pneumonia: a retrospective study. J Antimicrob Chemother 70(7): 2129–2132. doi: 10.1093/jac/dkv085. Epub 2015 Apr 21.
Sikkema RS, Farag EaBA, Himatt S, Ibrahim AK, Al-Romaihi H, Al-Marri SA, Al-Thani M, El-Sayed AM, Al-Hajri M, Haagmans BL, Koopmans MPG, Reusken CBEM (2017): Risk Factors for Primary Middle East Respiratory Syndrome Coronavirus Infection in Camel Workers in Qatar During 2013–2014: A Case-Control Study. J Infect Dis 215(11): 1702–1705. doi: 10.1093/infdis/jix174.
Simonsen L, Taylor RJ, Viboud C, Miller MA, Jackson LA (2007): Mortality benefits of influenza vaccination in elderly people: an ongoing controversy. Lancet Infect Dis 7(10): 658–666. doi: 10.1016/S1473-3099(07)70236-0.
Tripp RA, Haynes LM, Moore D, Anderson B, Tamin A, Harcourt BH, Jones LP, Yilla M, Babcock GJ, Greenough T, Ambrosino DM, Alvarez R, Callaway J, Cavitt S, Kamrud K, Alterson H, Smith J, Harcourt JL, Miao C, Razdan R, Comer JA, Rollin PE, Ksiazek TG, Sanchez A, Rota PA, Bellini WJ, Anderson LJ (2005): Monoclonal antibodies to SARS-associated coronavirus (SARS-CoV): identification of neutralizing and antibodies reactive to S, N, M and E viral proteins. J Virol Methods 128(1-2): 21–28. doi: 10.1016/j.jviromet.2005.03.021.
Van Doremalen N, Bushmaker T, Munster VJ (2013): Stability of Middle East respiratory syndrome coronavirus (MERS-CoV) under different environmental conditions. Euro Surveill 18(38): 20590. doi: 10.2807/1560-7917.es2013.18.38.20590.
Van Kerkhove MD, Alaswad S, Assiri A, Perera R, Peiris M, El Bushra HE, Binsaeed AA (2019): Transmissibility of MERS-CoV Infection in Closed Setting, Riyadh, Saudi Arabia, 2015. Emerg Infect Dis 25(10): 1802–1809. doi: 10.3201/eid2510.190130. Epub 2019 Oct 17.
Virlogeux V, Park M, Wu J, Cowling B (2016): Association between Severity of MERS-CoV Infection and Incubation Period. Emerg Infect Dis 22(3): 526–528. doi: 10.3201/eid2203.151437.
Wang J-T, Sheng W-H, Fang C-T, Chen Y-C, Wang J-L, Yu C-J, Chang S-C, Yang P-C (2004): Clinical manifestations, laboratory findings, and treatment outcomes of SARS patients. Emerg Infect Dis 10(5): 818–824. doi: 10.3201/eid1005.030640.
WHO (2012): https://www.who.int/csr/disease/coronavirus_infections/update_20121130/….
WHO (2019): https://www.who.int/emergencies/mers-cov/en/.
Widagdo W, Raj VS, Schipper D, Kolijn K, Van Leenders G, Bosch BJ, Bensaid A, Segalés J, Baumgärtner W, Osterhaus A, Koopmans MP, Van Den Brand JMA, Haagmans BL (2016): Differential Expression of the Middle East Respiratory Syndrome Coronavirus Receptor in the Upper Respiratory Tracts of Humans and Dromedary Camels. J Virol 90(9): 4838–4842. doi: 10.1128/JVI.02994-15. Print 2016 May.
Widagdo W, Sooksawasdi Na Ayudhya S, Hundie GB, Haagmans BL (2019): Host Determinants of MERS-CoV Transmission and Pathogenesis. Viruses 11(3): 280. doi: 10.3390/v11030280.
Yang Y, Zhang L, Geng H, Deng Y, Huang B, Guo Y, Zhao Z, Tan W (2013): The structural and accessory proteins M, ORF 4a, ORF 4b, and ORF 5 of Middle East respiratory syndrome coronavirus (MERS-CoV) are potent interferon antagonists. Protein Cell 4(12): 951–961. doi: 10.1007/s13238-013-3096-8. Epub 2013 Dec 8.
Yang Y, Ye F, Zhu N, Wang W, Deng Y, Zhao Z, Tan W (2015): Middle East respiratory syndrome coronavirus ORF4b protein inhibits type I interferon production through both cytoplasmic and nuclear targets. Sci Rep 5: 17554. doi: 10.1038/srep17554.
Zaki AM, Van Boheemen S, Bestebroer TM, Osterhaus ADME, Fouchier RaM (2012): Isolation of a Novel Coronavirus from a Man with Pneumonia in Saudi Arabia. N Engl J Med 367(19): 1814–1820. doi: 10.1056/NEJMoa1211721. Epub 2012 Oct 17.
Zhang W, Zheng XS, Agwanda B, Ommeh S, Zhao K, Lichoti J, Wang N, Chen J, Li B, Yang XL, Mani S, Ngeiywa KJ, Zhu Y, Hu B, Onyuok SO, Yan B, Anderson DE, Wang LF, Zhou P, Shi ZL (2019): Serological evidence of MERS-CoV and HKU8-related CoV co-infection in Kenyan camels. Emerg Microbes Infect 8(1): 1528–1534. doi: 10.1080/22221751.2019.1679610.
Zhao J, Perera RaPM, Kayali G, Meyerholz D, Perlman S, Peiris M (2015): Passive Immunotherapy with Dromedary Immune Serum in an Experimental Animal Model for Middle East Respiratory Syndrome Coronavirus Infection. J Virol 89(11): 6117–6120. doi: 10.1128/JVI.00446-15. Epub 2015 Mar 18.
Zielecki F, Weber M, Eickmann M, Spiegelberg L, Zaki AM, Matrosovich M, Becker S, Weber F (2013): Human cell tropism and innate immune system interactions of human respiratory coronavirus EMC compared to SARS-coronavirus. J Virol 87(9): 5300–5304. doi: 10.1128/JVI.03496-12. Epub 2013 Feb 28.
Zumla A, Hui DS, Al-Tawfiq JA, Gautret P, Mccloskey B, Memish ZA (2014): Emerging respiratory tract infections. Lancet Infect Dis 14(10): 910–911. doi: 10.1016/S1473-3099(14)70899-0. Epub 2014 Sep 1.
Zumla A, Hui DS, Perlman S (2015): Middle East respiratory syndrome. Lancet 386(9997): 995–1007. doi: 10.1016/S0140-6736(15)60454-8. Epub 2015 Jun 3.
Zumla A, Chan JFW, Azhar EI, Hui DSC, Yuen K-Y (2016): Coronaviruses – drug discovery and therapeutic options. Nat Rev Drug Discov 15(5): 327–347. doi: 10.1038/nrd.2015.37. Epub 2016 Feb 12.
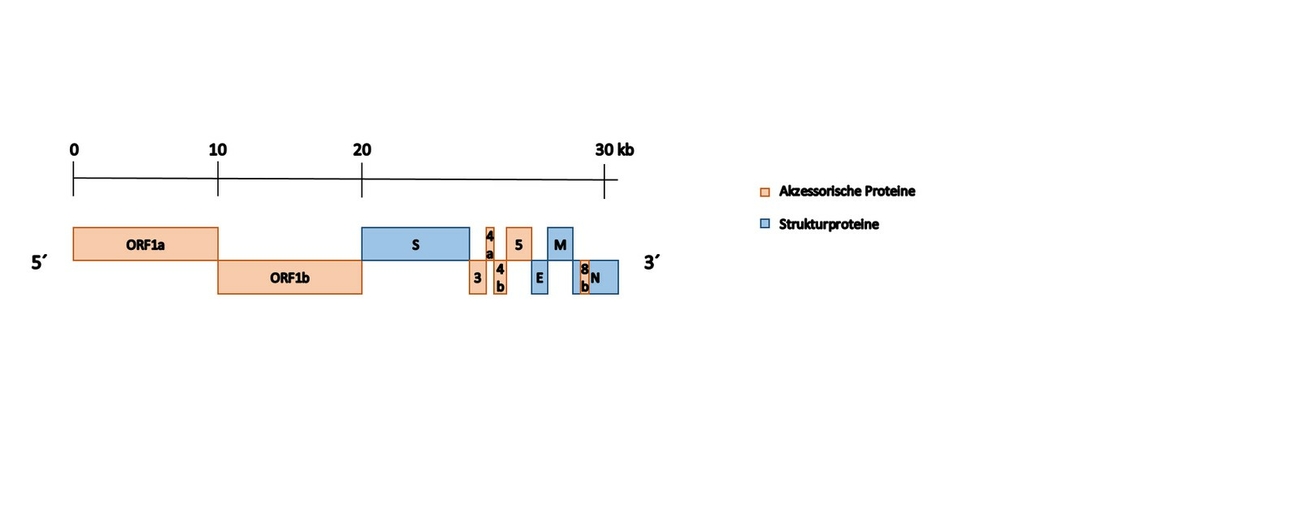


Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2020-30-Volz.pdf (0.23 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2020-30-Volz-Tab1.pdf (0.12 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2020-30-Volz-Tab2.pdf (0.19 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2020-30-Volz-Tab3.pdf (0.12 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2020-30-Volz-Tab4.pdf (0.19 MB) herunterladen möchten



{kind=link}
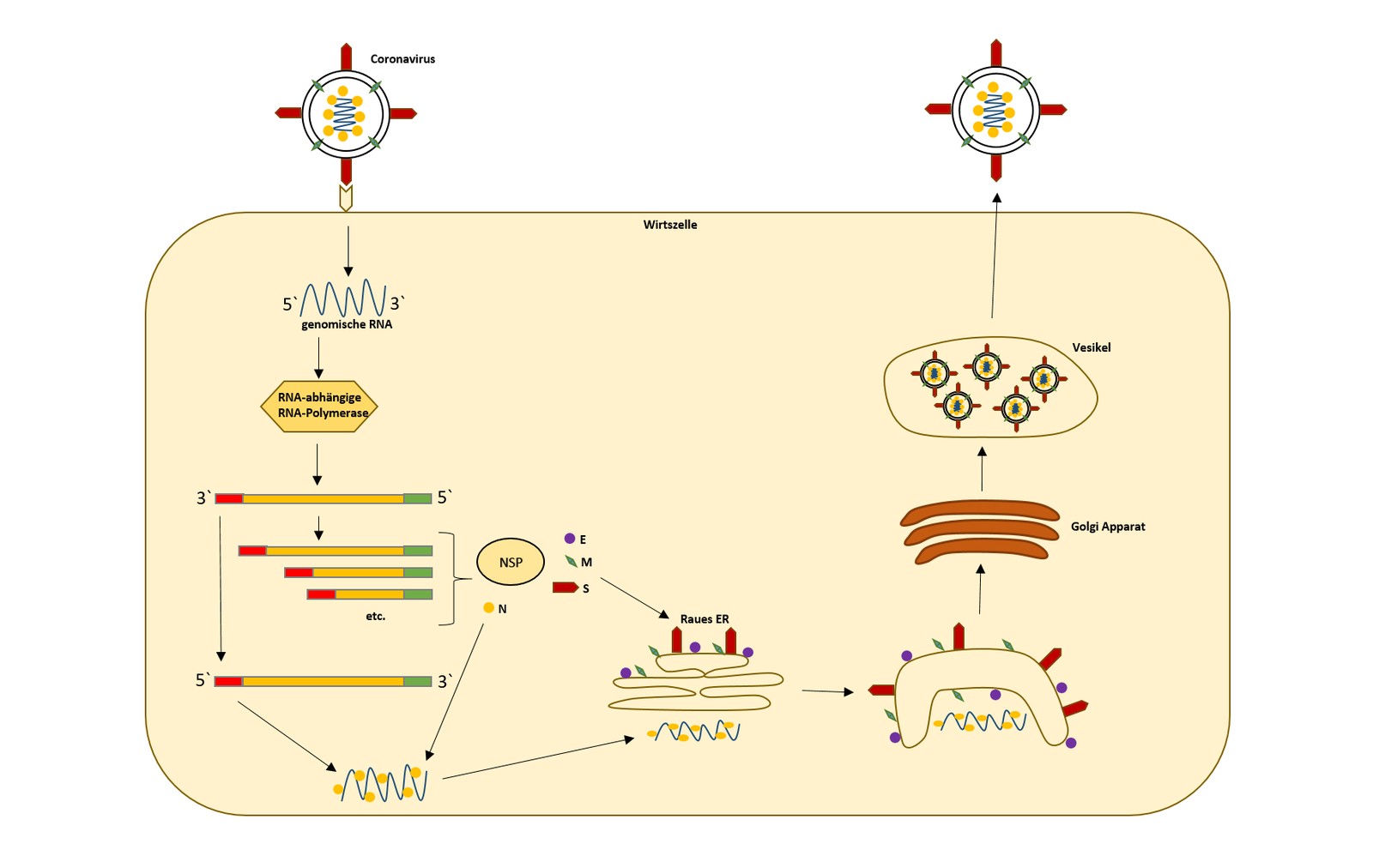{kind=link}