

Hämorrhagische Septikämie (Wild- und Rinderseuche) bei Rindern in Baden-Württemberg (Deutschland)
Berliner und Münchener Tierärztliche Wochenschrift 133
DOI: 10.2376/1439-0299-2020-8
© Schlütersche Verlagsgesellschaft mbH & Co. KG. 2020
Publiziert: 08/2020
Summary
Haemorrhagic septicaemia (septicaemic pasteurellosis) caused by Pasteurella (P.) multocida capsular type B has been diagnosed in cattle in the federal state of Baden-Wuerttemberg (Germany) for the first time in June 2019 since its last official report in Germany in 1986 and its reoccurrence in 2010. A total of 13 cattle succumbed to sudden death on four farms located in the northern part of the Black Forest between June and November 2019. Post-mortem examinations were carried out on seven of these animals and all displayed marked hyperaemia, haemorrhages and oedema in the subcutis and in several inner organs. The pathological-anatomical findings were confirmed by histopathological analyses.
Cultivation of P. multocida was successful in five of seven cattle with high bacterial loads in internal organs, and poor growth in two animals. All isolates were clearly identified as P. multocida by MALDI-TOF mass spectrometry and were compared with isolates originating from an HS outbreak in eastern Germany in summer 2010 using Fourier transform infrared (FT-IR) spectroscopy. Molecular capsular and LPS genotyping assigned six of the isolates to the genotypes B:L2 and one to B:L6, respectively. All isolates belonged to the RIRDC MLST genotype ST122. Antimicrobial testing of P. multocida isolates originating from each farm based on the determination of minimal inhibitory concentration (MIC) revealed susceptibility to ampicillin, ceftiofur, enrofloxacin, florfenicol, penicillin G, tetracycline, and tulathromycin. Resistance was determined for spectinomycin.
Zusammenfassung
Die durch Pasteurella (P.) multocida Kapseltyp B verursachte Hämorrhagische Septikämie (Wild- und Rinderseuche) wurde bei Rindern zum ersten Mal in Baden-Württemberg Ende Juni 2019 seit der letzten offiziellen Meldung 1986 und dem Wiederauftreten in Deutschland im Jahr 2010 nachgewiesen. Insgesamt kam es zum perakuten Verenden von 13 Rindern in vier Betrieben in der Region des Nördlichen Schwarzwaldes zwischen Juni und November 2019. Pathologisch-anatomische Untersuchungen von insgesamt sieben Tieren zeigten Hyperämie, Hämorrhagien und Ödeme in der Unterhaut und verschiedenen inneren Organen in unterschiedlicher Ausprägung. Die pathologisch-anatomischen Veränderungen wurden durch pathohistologische Untersuchungen bestätigt.
Die Kultivierung des Erregers P. multocida gelang bei fünf von sieben Rindern mit starkem und bei zwei Tieren nur mit schwachem Wachstum in den inneren Organen. Alle Isolate wurden eindeutig mittels MALDI-TOF Massenspektrometrie als P. multocida identifiziert und mit Hilfe der Fourier-Transformations-Infrarot- (FT-IR) Spektroskopie mit Isolaten, die aus einem HS-Ausbruch in Ostdeutschland im Sommer 2010 stammten, verglichen. Molekulare Kapsel- und LPS-Genotypisierungen ermöglichten eine Zuordnung zum Genotyp B:L2 bei sechs und zum Genotyp B:L6 bei einem Isolat. Alle Isolate konnten dem RIRDC MLST Genotyp ST122 zugeordnet werden. Die Testung der P. multocida-Isolate gegen verschiedene Antibiotika ergab Empfindlichkeit gegen Ampicillin, Ceftiofur, Enrofloxacin, Florfenicol, Penicillin G, Tetracyclin und Tulathromycin. Spectinomycin erwies sich als resistent.
Introduction
Haemorrhagic septicaemia (HS) is a serious disease in cattle and other bovines. It is characterised by an acute and septicaemic course resulting in high mortality rates (OIE 2013, Shivachandra et al. 2011). HS in cattle is a disease listed by the OIE (OIE 2020). However, small ruminants and pigs, and horses and donkeys to a much lower extent, are also susceptible to this disease (De Alwis 1999). HS has also been recorded in wild ungulates (Carrigan et al. 1991, De Alwis 1999), and a devastating HS outbreak led to a mass extinction of about 200,000 critically endangered saiga antelopes in Kazakhstan in 2015 (Fereidouni et al. 2019).
The causative agent of HS is Pasteurella (P.) multocida, a gram negative, facultative anaerobic, non-motile bacterium. P. multocida isolates can be classified by capsular serotyping or by DNA-based capsular typing into the five groups A, B, D, E and F (Townsend et al. 2001) and further subtyped based on eight LPS genotypes (L1–L8) (Harper et al. 2015) comprising 16 serovars (Brogden et al. 1978, Heddleston et al. 1972). However, the occurrence of capsular non-typeable P. multocida isolates has to be considered (Arumugam et al. 2011). Usually, the capsular type B is found in HS cases in Asia and Europe (Asian capsular type) whereas type E is encountered in Africa (African capsular type) (De Alwis 1992). The occurrence of HS is usually climate-related and areas regularly hit are Asia, sub-Saharan Africa and southern Europe (Shivachandra et al. 2011). However, HS has also been reported in cattle, pigs and wildlife within the last decade in countries located in central Europe in temperate climate zones such as Hungary (Magyar et al. 2017, Ujvari et al. 2015), Spain (Risco et al. 2013), Denmark (Eriksen et al. 1999) and Germany (Falkenberg 2016, Kutzer et al. 2019, LUA 2019, Müller and Locher, 2017, Rohkohl et al. 2015, Soike et al. 2012, TSK Sachsen 2018, Völker et al. 2014). In Germany HS has reoccurred in 2010 (Soike et al. 2012) since the last officially reported cases in 1986 (OIE 2016, WAHIS 2019).
HS-causing isolates of P. multocida persist in the nasopharynx and tonsils of carrier animals (De Alwis et al. 1990, OIE 2013, Soike et al. 2012) or in the oral flora of wolves (Kutzer et al. 2019). Stressors such as high temperatures, lack of food and water or crowding promote immunosuppression and may thus trigger outbreaks of HS (Annas et al. 2014, De Alwis 1992, Soike et al. 2012).Transmission of the pathogen occurs by the nasal or oral route via inhalation or ingestion through direct contact with infected animals or contaminated fomites (Spickler 2009). Infected cattle develop an acute or even peracute course of infection with symptoms of septicaemia including fever, malaise, recumbency, dyspnoea, nasal discharge, oedema and haemorrhages resulting in high mortality rates. Post-mortem examinations reveal haemorrhages, oedema and hyperaemia in tissues and organs in accordance with the severity of the disease (OIE 2013, Shivachandra et al. 2011).
Top Job:
Starting with cases of HS in Brandenburg and Saxony-Anhalt (2010) (Soike et al. 2012), repeated limited outbreaks in several federal states during the last decade have led to the re-emergence of HS in Germany (Kutzer et al. 2019, Müller and Locher 2017, Rohkohl et al. 2015, TSK Sachsen 2018, Völker et al. 2014).
The present report describes the first cases of HS in cattle in the federal state of Baden-Wuerttemberg (Germany) between June and November 2019.
Material and Methods
General data on the cases
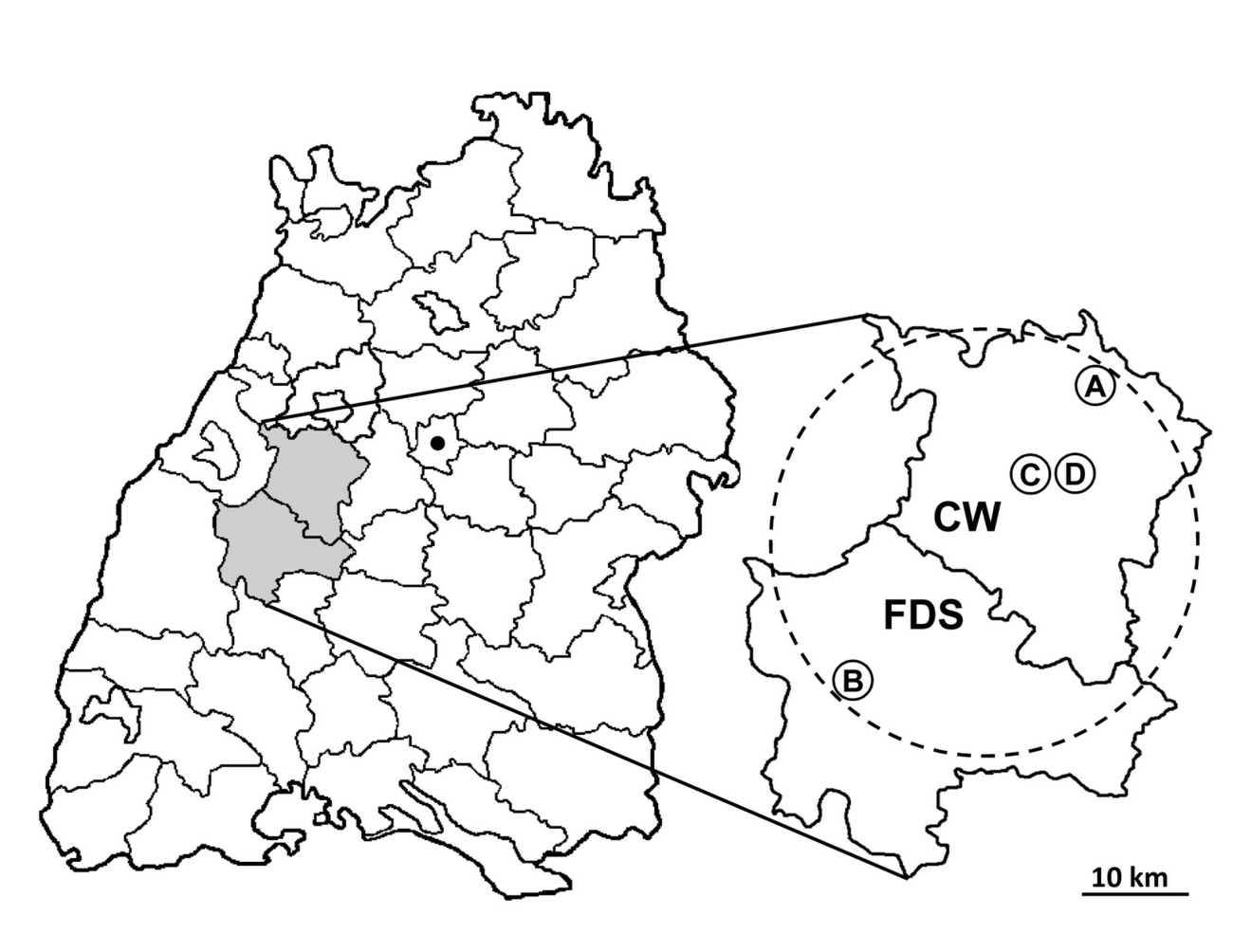
General data on the affected farms including the date of first registered HS cases, the herd localities and sizes, and the number, age and sex of the perished animals were collected. Post-mortem examinations were performed on seven cattle originating from four herds located in three municipalities from two districts (Table 1, Fig. 1). Data on the lowest and highest maximal outdoor temperatures per day for each month during the period in which the cases of HS occurred was accessed from the open access database (www.wetterkontor.de).
Pathological examinations
The carcasses were subjected to pathological-anatomical and histopathological examinations. Tissue samples from the organs showing gross lesions were submitted for histological analyses using haematoxylin-eosin staining.
Microbiological examinations
For cultivation of bacteria, aseptically obtained specimens from lung, liver, spleen, kidney, mesenteric lymph nodes and other visibly affected organs were streaked directly onto 5% sheep blood agar and water-blue metachrome-yellow lactose agar according to Gassner. In addition, specimens of the lungs were smeared on Pasteurella Selective Agar (Oxoid, Wesel, Germany). The agar plates were incubated at 37°C under aerobic conditions for at least 48 h. For further analyses, colonies indicative for Pasteurella species were sub-cultured on agar with 5 % sheep blood to obtain pure cultures. The MALDI-TOF mass-spectra generated by the microflex LT System (Bruker Daltonik, Bremen, Germany) were analysed using the Bruker Biotyper software Version 3.1 and evaluated with the Bruker Taxonomy database (DB 8.468 entries, Bruker Daltonik). Analyses were extended with an in-house database, including a set of 13 reference spectra of known P. multocida capsule types (MUP 0924–0931, 0933, 0934, 0937, 0942, 1503), comprising representatives of capA, capD, capF, capsular non-typeable, and five reference spectra of P. multocida isolates from capsular type B or E (Rau et al. 2016a). The creation of new reference entries, so-called main spectra projections (MSP), followed the manufacturer’s instructions and standards (Pranada et al. 2016, Rau et al. 2016b).
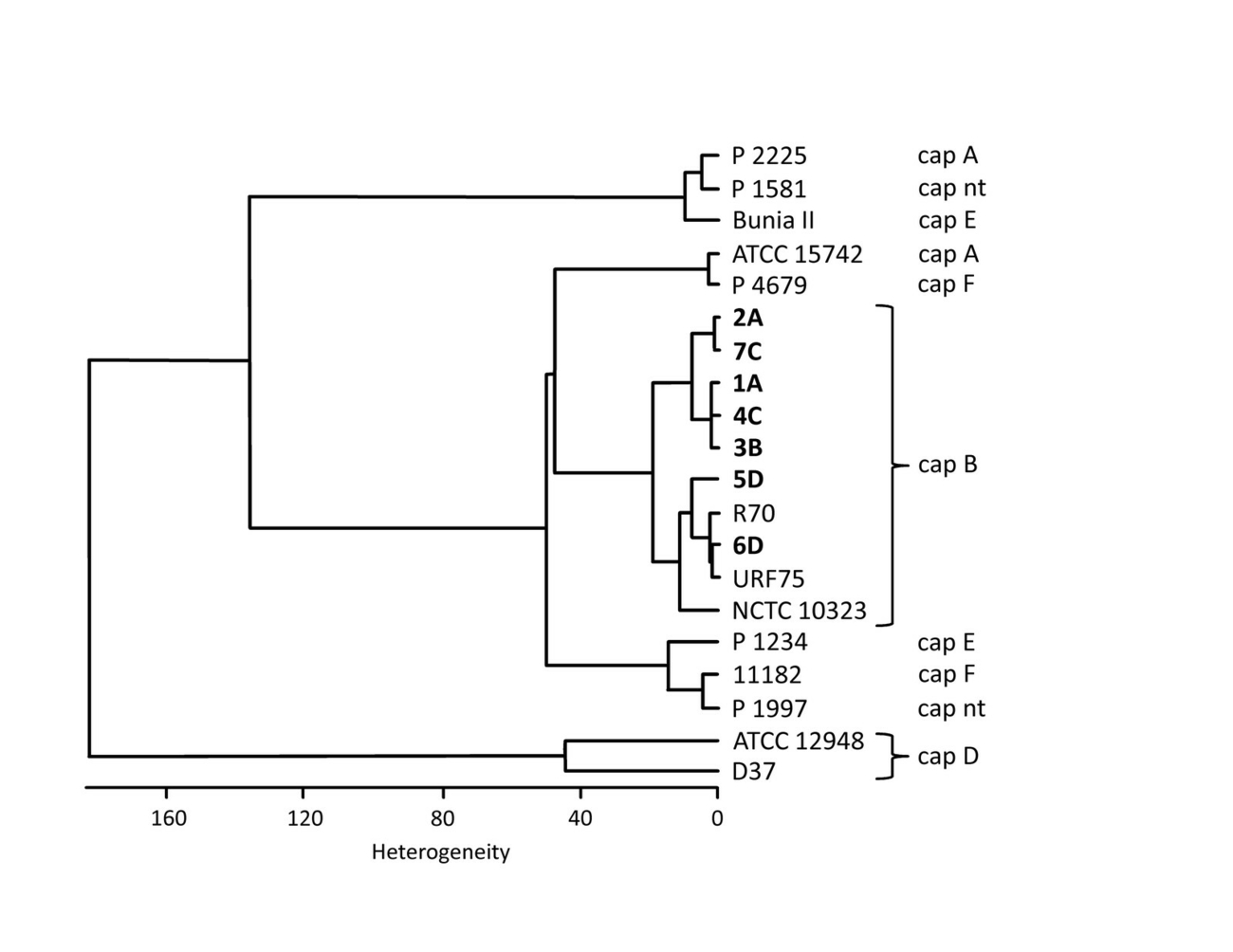
FT-IR (Fourier transform infrared) spectroscopy analyses were carried out and evaluated with a Tensor 27 spectrometer equipped with an HXT module (Bruker Optics, Ettlingen, Germany) (Contzen et al. 2011). Cluster analysis (Ward’s algorithm) was performed using the second derivatives of the spectra in the spectral range 500–1450 cm-1. The IR-spectra of the isolates were compared with a collection of P. multocida including several capsular type B isolates (Fig. 2).
For the identification of HS-causing P. multocida isolates we used the OIE-recommended HS-causing-type-B-specific PCR according to Townsend et al. (1998). LPS genotyping was carried out using the multiplex PCR described by Harper et al. (2015). Furthermore, all isolates were subjected to the RIRDC MLST scheme developed for P. multocida based on PCR amplification and sequencing of seven housekeeping genes according to Subaaharan et al. (2010). The sequence types were determined with help of the Pasteurella multocida MLST Databases of PUBMLST (https://pubmlst.org/pmultocida/) (Jolley et al. 2004). For this purpose, genomic DNA was extracted using the QIAGEN Genomic-tip 100/G kit (Qiagen, Hilden, Germany) and the Roche High Pure PCR Template Preparation Kit (Roche, Mannheim, Germany) according to the manufacturers’ instructions from a single colony grown in LB medium supplemented with 1% glucose at 37°C.
Minimum inhibitory concentrations (MIC) of antimicrobial drugs for P. multocida isolates recovered from the cattle 2, 3, 4, and 5 (Table 1) were determined based on the microdilution method using the antibiotic microdilution plate Micronaut-S Veterinary Large Animal (MERLIN, Bornheim-Hersel, Germany). Antimicrobial susceptibility testing was carried out according to the manufacturer’s instructions and evaluated according to the CLSI guidelines (CLSI 2018) (Table 3).
Results
General data on the cases
A total of 13 cattle succumbed to sudden death on four farms in the federal state of Baden-Wuerttemberg (Germany). The farms are located in the administrative districts of Calw and Freudenstadt in the northern Black Forest region (Fig. 1). The HS cases occurred within a period of approx. four months (June/July to November 2019). Eleven cattle died on the farms’ pastures and two cows in an open loose-housing barn. From all seven cattle submitted for post-mortem examinations, P. multocida could be cultivated (Table 1). Antibiotics were administered only in farm A, starting with enrofloxacin for two days and continued with penicillin for further three days. The minimum distance between the farms is 2 km and they are located within a circle with a diameter of 40 km (Fig. 1). The animals on these farms had neither direct nor indirect contact. The cattle grazed in groups of 30 (farm A), 14 (farm B), and 28 (farm C) individuals, respectively. In farm D 164 cows lived in an open loose-housing barn. All farms are located directly at forest borders of the eastern side of the northern Black Forest and are separated by large areas of forest and meadows which have been partly visited by wild animals such as wild boars.
All animals submitted to post-mortem examinations had been found dead from one day to the next without previous clinical signs such as fever, recumbency, stupor, malaise or dyspnoea. Details on the animals subjected to post-mortem examinations, and details on animals that died peracutely on these farms but were not examined are listed in Table 1.
Analyses of the climate data for the northern Black Forest (weather station Neubulach near Bad Teinach-Zavelstein, district of Calw) during the period of the HS outbreaks revealed maximum temperatures per day, as follows: June 15.6–33.8 °C, July 18.0–34.2 °C, August 16.8–31.0 °C, September 11.7–27.4 °C, October 6.3–24.4 °C, and November (until 15. November) 2.5–15.9 °C.
Pathology
On first inspection, all animals were in good physical and nutritional condition. Post-mortem examinations revealed hyperaemia, haemorrhage or oedema in various organs or in at least one organ with different severity. The kidneys of all seven animals did not display visible pathological changes. The pathological findings are summarised in Table 2.
The pathological anatomical changes were corroborated by pathohistological investigations. Examination of six cattle revealed hyperaemia, haemorrhages and oedema in the lung (cattle 2, 3 and 7), liver (cattle 2), kidney (cattle 5), meninges (cattle 1, 2, 6), udder (cattle 1 and 2), heart (cattle 1), skeletal muscles (cattle 1) or subcutis (cattle 2) at different intensities. In addition, necrosis of the renal tubules (cattle 2), lungs and muscles (cattle 3) were found. In cattle 4, interstitial oedema and necrotising vasculitis in the tissue between oesophagus, trachea and buccal mucosa was apparent. Cattle 5 showed scarring and interstitial fibrosis of the lung.
Microbiological examinations
High bacterial load of P. multocida was detected in lung, liver, spleen, kidney and lymph nodes in animals 1 to 4 and 7 after cultivation on blood agar for 24 h. Due to the pathological findings, swabs were also taken from the pleura, urinary bladder, udder and muscle (cattle 1) and from the brain, heart, subcutis and udder (cattle 2), from muscle (cattle 3), and from blood and subcutis (cattle 7), respectively. These specimens also showed very strong P. multocida bacterial growth.
In cow 5, only two colonies of P. multocida could be cultivated from the liver and the spleen, respectively and in cattle 6, cultivation of P. multocida was only successful from the spleen and the pleura with poor and extensive bacterial growth, respectively.
In antimicrobial susceptibility testing, P. multocida isolates originating from all four farms were sensitive to ampicillin, ceftiofur, enrofloxacin, florfenicol, penicillin G, tetracycline, and tulathromycin and resistant to spectinomycin (Table 3).
MALDI-TOF MS
The putative P. multocida isolates were confirmed at species level by MALDI-TOF MS with the commercial Bruker database (score values 2.315 ± 0,051). The application of the database, which had been extended by 13 additional reference entries for capsular type defined strains, resulted in an increase of the score values to 2.619 ± 0.065. The hit lists for the isolates from the HS cases were headed by matches of P. multocida from both HS causing capsule types B and E.
Capsular typing
Fourier-transform infrared (FT-IR) spectroscopy with cluster analysis was carried out as described previously (Nagib et al. 2014). The available isolates from HS cases described in this study were compared with isolates from HS cases from eastern Germany in 2010/2011, as well as other P. multocida with defined capsule types (Soike et al. 2012). The cluster diagram shows the level of uniformity between the capB-type P. multocida from the 2019 event, and the similarity to isolates (R70, URF75) from the HS cases 2010/2011 from Germany (Fig. 2).
The type-B-specific PCR for HS-causing P. multocida bacteria consistently retrieved a positive result. Additional testing on the capsular type confirmed that all isolates belong to capsular type B.
LPS genotyping allocated the isolates obtained from the animals 1–4 and 6–8 to genotype L2, which includes the Heddleston serovars 2 and 5. LPS genotype L6 including the Heddleston serovars 10, 11, 12 and 15 was determined for animal 5.
Using RIRDC MLST all isolates could be assigned to the clonal group ST122.
Discussion
Clinical signs, pathology
The cases of HS in cattle described in this report developed typical acute or peracute, predominantly sudden fatal outcomes of the disease without obvious or only subtle clinical signs the day before. Pathological changes were characterised by hyperaemia, haemorrhage and oedema in several organs. The major inner organs lung, liver, spleen and the serosa of the body cavities were most strongly affected by hyperaemia and haemorrhages. Additionally, the brain, udder and muscles were also visibly affected in some cases. Oedemata were present in the lung and subcutis. These pathological findings were pronounced at different intensities and could be confirmed by histopathological examinations. Völker et al. (2014) also outline comparable pathological lesions and Soike et al. (2012) comprehensively described pathological findings in cattle, fallow dear and pigs that had succumbed to death due to recent outbreaks of HS. Severe organ lesions are caused by virulence factors of P. multocida capsular type B, especially endotoxins (LPS) triggering a sudden and immense secretion of proinflammatory cytokines provoking endotoxic shock symptoms and finally leading to death (Harper and Boyce 2017, Horadagoda et al. 2001).
The pathogen
In the cases reported here, HS led to a high bacterial load of P. multocida in the lung, liver, spleen and kidney in five of seven examined cattle. The affected organs, i.e. brain, udder, muscle, heart and the serosa of body cavities corresponded with those recently described by Völker et al. (2014) and Soike et al. (2012). In two cases, however, P. multocida was detected in two organs only (kidney and spleen, spleen and pleura), although the animals had not been treated with antibiotics. With the exception of the pleura, the bacterial load was low. This observation raises the question of the role of bacterial load, virulence associated genes and pathogenesis of P. multocida during HS (Verma et al. 2013).
P. multocida was identified unambiguously using MALDI-TOF MS and the capsular type was determined by FT-IR analyses and verified by capsular-PCR (Soike et al. 2012, Townsend et al. 2001).
The FT-IR cluster diagram shows a high level of uniformity between the seven capB-type P. multocida from the 2019 event, and a great similarity to isolates from the HS cases 2010/2011 from Germany (Fig. 2).
Further LPS genotyping revealed HS isolates of the genotype L2 (comprising serovars 2, 5) in all farms. However, a P. multocida isolate of the LPS genotype L6 (comprising the serovars 10–12, 15) was isolated from an animal on farm A and a day later an isolate with the LPS genotype L2 from another animal on the same farm. It has been reported that HS in cattle and buffalo is exclusively caused by the LPS genotype 2 (Harper and Boyce 2017). Even though, P. multocida belonging to the LPS genotype 6 had been retested with the same result and was unequivocally associated with pathological anatomical findings typical for HS in cattle this finding remains unclear. However, all except for one isolate could be assigned to the genotype B:L2:ST122 and all isolates to the clonal MLST group ST122. This is in agreement with findings for isolates associated with HS in bovines originating from Europe (Hotchkiss et al. 2011, Soike et al. 2012) and for isolates which been collected in Asia and India (Hotchkiss et al. 2011, Peng et al. 2018, Petersen et al. 2014).
Testing of antimicrobial susceptibility revealed sensitivity to a broad spectrum of antibiotics, including ampicillin, ceftiofur , enrofloxacin, florfenicol, penicillin G, tetracycline, and tulathromycin for all P. multocida isolates. These results are in agreement with published data (BVL 2019, OIE 2018, Rohkohl et al. 2015, Soike et al. 2012).
Epidemiology
In Germany, the last cases of HS were officially reported in 1986 (OIE 2016, WAHIS 2019). However, the re-emergence of HS in Germany has been confirmed by several outbreaks over the past decade. Cases have been reported in Brandenburg and Saxony-Anhalt (2010) (Soike et al. 2012), Lower Saxony (2013/2014) (Rohkohl et al. 2015, Völker et al. 2014), Mecklenburg-Western Pomerania (2014) (Falkenberg 2016), and recently in Bavaria (Müller and Locher 2017), Saxony (TSK Sachsen 2018), Thuringia (Kutzer et al. 2019), Rhineland-Palatinate (LUA 2019), and in Baden-Wuerttemberg (present study). All outbreaks in cattle have in common that animal deaths occurred mainly during the hot season in association with grazing. In the present report, the temperature exceeded 30°C in June. The first HS cases occurred within one week and further sporadic cases followed in the subsequent three months. The last case of HS was in late November when the temperatures had already dropped below 16°C. HS outbreaks regardless of the season have been reported in subtropical (India) (Kumar et al. 2004) and temperate regions (Germany) (Rohkohl et al. 2015). The occurrence of HS at various locations within a 50 km diameter and a time period of only one week has been reported by Rohkohl et al. (2015) and Soike et al. (2012). Rohkohl et al. (2015) described the onset of an HS outbreak in the warm season and recurrence in November due to latently infected cows returning into the cowshed. This phenomenon can be explained by persistence of the pathogen in the crypts of tonsils and low infectious doses of virulent bacteria (Shivachnadra et al. 2011, Soike et al. 2012). What causes the pathological findings despite low bacterial load in the inner organs is still unknown, as are the pathomechanisms and pathogenesis of septicaemic P. multocida infections.
To date, the source of HS and the pathogen spread over long distances, especially to southwestern Germany, remains unresolved. Possible explanations for these outbreaks are direct or indirect contact with wildlife, which could function as carrier (Kutzer et al. 2019, Soike et al. 2012). However, the proof of the source, spread and transmission of infections to livestock is also still pending for the cases reported for Germany (Rohkohl et al. 2015, Soike et al. 2012, Völker et al. 2014).
Conclusion
HS has re-emerged in Germany in the past decade. The source of infection, the reservoir, the mode of transmission and the pathomechanisms are still poorly understood. Farmers, veterinarians and hunters should be alerted by sudden cases of deaths in domestic and feral even-toed ungulates. In cases of HS outbreaks, immediate removal of dead animals, implementation of the principles of ‘stand still’, and metaphylactic administration of antibiotics in the short term and vaccination with adequate LPS serotypes in the long term represent effective measures. To prevent outbreaks of HS, it is essential to know the extent and spread of this disease and the sources of infection. Further studies are urgently needed to increase knowledge about HS in central Europe.
The exchange of reference spectra of capsule type defined P. multocida isolates is facilitated by the MALDI-user platform (https://maldi-up.ua-bw.de).
Acknowledgement
The authors thank Sandra Ketterer-Pintur and Brigitte Illi for evaluation of the bacterial cultures, Martin Dyk for the MALDI-TOF MS and FT-IR analysis, and Maren Münch for performance and evaluation of the antimicrobial resistance tests.
Conflict of interest
The authors declare that they have no conflicts of interest.
Ethical approval
Since this is a study on fatal cases of cattle and no living animals were involved, no ethical vote was necessary. The authors declare that no research interventions or experiment with animals or human beings, no clinical trials or clinical research were conducted in the context of this study.
Funding
The authors declare that they did not receive any financial support for this publication.
Authors contribution
RS: Project idea, planning and conception of this study, literature research, writing and revision of the manuscript, data analysis.
BB, IS and AKS: Performance of the pathological-anatomical examinations and the histopathological analysis.
SM: Collection and analysis of the field data.
LDS: Performance and analysis of the molecular investigations.
JR: Performance and evaluation of the MALDI-TOF MS and FT-IR studies.
All authors contributed to this manuscript by writing and revision and approved the final version.
Address for correspondence
Dr. Reinhard Sting
Chemisches und Veterinäruntersuchungsamt Stuttgart
Schaflandstr. 3/3
70736 Fellbach
Germany
reinhard.sting@cvuas.bwl.de
References
Annas S, Zamri-Saad M, Jesse FF, Zunita Z (2014): New sites of localisation of Pasteurella multocida B:2 in buffalo surviving experimental haemorrhagic septicaemia. BMC Vet Res 10: 88.
Arumugam ND, Ajam N, Blackall PJ, Asiah NM, Ramlan M, Maria J, Yuslan S, Thong KL (2011): Capsular serotyping of Pasteurella multocida from various animal hosts – a comparison of phenotypic and genotypic methods. Trop Biomed 28: 55–63.
Brogden KA, Rhoades KR, Heddleston KL (1978): A new serotype of Pasteurella multocida associated with fowl cholera. Avian Dis 22: 185–190.
BVL (2019): Bericht zur Resistenzmonitoringstudie 2017. BVL-Report 13.7. Bundesamt für Verbraucherschutz und Lebensmittelsicherheit (BVL). https://www.bvl.bund.de/DE/Arbeitsbereiche/09_Untersuchungen/01_Aufgabe….
Carrigan MJ, Dawkins HJ, Cockram FA, Hansen AT (1991): Pasteurella multocida septicaemia in fallow deer (Dama dama). Aust Vet J 68: 201–203.
CLSI (2018): Performance standards for antimicrobial disc and dilution susceptibility tests for bacteria isolated from animals. 4th ed. CLSI supplement VET08. Wayne, PA: Clinical and Laboratory Standards Institute.
Contzen M, Sting R, Blazey B, Rau J (2011): Corynebacterium ulcerans from diseased wild boars. Zoonoses Public Health 58: 479–488.
De Alwis MCL (1992): Haemorrhagic septicaemia – a general review. Br Vet J 148: 99–112.
De Alwis MCL (1999): Haemorrhagic septicaemia. ACIAR Monograph No. 57, Canberra, Australia.
De Alwis MCL, Wijewardana TG, Gomis AIU, Vipulasiri AA (1990): Persistence of the carrier status in haemorrhagic septicaemia (Pasteurella multocida serotype 6:B infection) in buffalo. Trop Anim Health Pro 22: 185–194.
Eriksen L, Aalbaek B, Leifsson PS, Basse A, Christiansen T, Eriksen E, Rimler RB (1999): Hemorrhagic septicemia in fallow deer (Dama dama) caused by Pasteurella multocida multocida. J Zoo Wildl Med 30: 285–292.
Falkenberg U (2016): „Pasteurellen einmal anders“ – Die Hämorrhagische Septikämie oder Wild- und Rinderseuche in Rinderbeständen in MV. Veranstaltung des bpt Landesverbandes MV und LTÄK MV am 19.10.2016 in Güstrow. www.bpt-mv.de/Praxis-Service/UFalkenberg_Gustrow-offentlich.pdf.
Fereidouni S, Freimanis GL, Orynbayev M, Ribeca P, Flannery J, King DP, Zuther S, Beer M, Höper D, Kydyrmanov A, Karamendin K, Kock R (2019): Mass die-off of Saiga antelopes, Kazakhstan, 2015. Emerg Infect Dis 25: 1169–1176.
Harper M, Boyce JD (2017): The myriad properties of Pasteurella multocida lipopolysaccharide. Toxins 9: 254.
Harper M, John M, Turni C, Edmunds M, St Michael F, Adler B, Blackall PJ, Cox AD, Boyce JD (2015): Development of a rapid multiplex PCR assay to genotype Pasteurella multocida strains by use of the lipopolysaccharide outer core biosynthesis locus. J Clin Microbiol 53: 477–485.
Heddleston KL, Gallagher JE, Rebers PA (1972): Fowl cholera: Gel diffusion precipitin test for serotyping Pasteurella multocida from avian species. Avian Dis 16: 925–936.
Horadagoda NU, Hodgson JC, Moon GM, Wijewardana TG, Eckersall PD (2001): Role of endotoxin in the pathogenesis of haemorrhagic septicaemia in the buffalo. Microb Pathog 30: 171–178.
Hotchkiss EJ, Hodgson JC, Lainson FA, Zadoks RN (2011): Multilocus sequence typing of a global collection of Pasteurella multocida isolates from cattle and other host species demonstrates niche association. BMC Microbiol 11: 115.
Jolley KA, Chan MS, Maiden MCJ (2004): mlstdbNet – distributed multi-locus sequence typing (MLST) databases. BMC Bioinform 5: 86.
Kumar AA, Shivachandra SB, Biswas A, Singh VP, Singh VP, Srivastava SK (2004): Prevalent serotypes of Pasteurella multocida isolated from different animal and avian species in India. Vet Res Commun 28: 657–667.
Kutzer P, Szentiks C, Bock S, Ewers C, Schulze C, Semmler T (2019): Die Rückkehr der Hämorrhagischen Septikämie: Der Wolf als Vektor? 38. Jahrestagung der DVG-Fachgruppe AVID – Schwerpunkt Bakteriologie, Staffelstein 11.–13.09.2019.
LUA (Landesuntersuchungsamt) Rheinland-Pfalz (2019): Wild- und Rinderseuche: Erster Nachweis in Rheinland-Pfalz. Tierseuchen 22.08.2019. https://lua.rlp.de/de/presse/detail/news/News/detail/wild-und-rinderseu….
Magyar T, Ujvári B, Szeredi L, Virsinger N, Albert E, Német Z, Csuka E, Biksi I (2017): Re-emergence of bovine haemorrhagic septicaemia in Hungary. Acta Vet Hung 65: 41–49.
Müller M, Locher N (2017): Hämorrhagische Septikämie bei Wildtieren und Rindern in Nordbayern. Bayerisches Landesamt für Gesundheit und Lebensmittelsicherheit. https://www.lgl.bayern.de/tiergesundheit/tierkrankheiten/bakterielle_pi….
Nagib S, Rau J, Sammra O, Lämmler C, Schlez K, Zschöck M, Prenger-Berninghoff E, Klein G, Abdulmawjood A (2014): Identification of Trueperella pyogenes isolated from bovine mastitis by Fourier Transform Infrared Spectroscopy. PLoS One 9: e104654.
OIE (World Organisation for Animal Health) (2013): Haemorrhagic Septicaemia. OIE Technical Disease Card. https://www.oie.int/fileadmin/Home/eng/Animal_Health_in_the_World/docs/….
OIE (World Organisation for Animal Health) (2016): Handistatus II, Last update 14. December 2016. http://web.oie.int/hs2/report.asp.
OIE (World Organisation for Animal Health) (2018): Haemorrhagic septicaemia. Chapter 3.4.10. https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.04.10_HA….
OIE (World Organisation for Animal Health) (2020): OIE-Listed diseases, infections and infestations in force in 2020. www.oie.int/en/animal-health-in-the-world/oie-listed-diseases-2020/.
Peng Z, Liang W, Wang F, Xu Z, Xie Z, Lian Z, Hua L, Zhou R, Chen H, Wu B (2018): Genetic and phylogenetic characteristics of Pasteurella multocida isolates from different host species. Front Microbiol 9: 1408.
Petersen A, Bisgaard M , Townsend K, Henrik Christensen H (2014): MLST typing of Pasteurella multocida associated with haemorrhagic septicaemia and development of a real-time PCR specific for haemorrhagic septicaemia associated isolates. Vet Microbiol 170: 335–341.
Pranada, BP, Schwarz G, Kostrzewa M (2016): MALDI biotyping for microorganism identification in clinical microbiology. In: Cramer R (ed.), Advances in MALDI and laser-induced soft ionization mass spectrometry. Springer International Publishing, Basel, 197–226.
Rau J, Männig A, Hiller E, Mauder N, Wind C, Horlacher S, Kadlec K, Schwarz S, Contzen M (2016a): MALDI-TOF mass spectrometry for reliable identification of 444 bacteria – A validation based on Staphylococcacae field isolates. Aspects of Food Control and Animal Health 03/2016, 446. https://ejournal.cvuas.de/issue201603.asp.
Rau J, Eisenberg T, Sting R (2016b): MALDI-UP – an internet platform for the exchange of MALDI-TOF mass spectra.
https://maldi-up.ua-bw.de.
Risco D, Fernández-Llario P, Cuesta JM, García-Jiménez WL, Gil M, Gonçalves P, Martínez R, Gómez L, García A, Rey J, Hermoso de Mendoza M, Hermoso de Mendoza JH (2013): Fatal outbreak of systemic pasteurellosis in a wild boar (Sus scrofa) population from southwest Spain. J Vet Diagn Invest 25: 791–794.
Rohkohl J, Schulze C, Bilk S (2015): Hämorrhagische Septikämie bei Milchkühen in Niedersachsen. Prakt Tierarzt 96: 598–608.
Shivachandra SB, Viswas KN, Kumar AA (2011): A review of hemorrhagic septicemia in cattle and buffalo. Anim Health Res Rev 12: 67–82.
Soike D, Schulze C, Kutzer P, Ewert B, van der Grinten E, Schliephake A, Ewers C, Bethe A, Rau J (2012): Acute pasteurellosis in fallow deer, cattle and pigs in a region of Eastern Germany. Berl Munch Tierarztl Wochenschr 125: 122–128.
Spickler AR (2009): Hemorrhagic septicemia. Fact sheet. Center for Food Security and Public Health (CFSPH). http://www.cfsph.iastate.edu/Factsheets/pdfs/hemorrhagic_septicemia.pdf.
Subaaharan S, Blackall LL, Blackall PJ (2010): Development of a multi-locus sequence typing scheme for avian isolates of Pasteurella multocida. Vet Microbiol 141: 354–361.
Townsend KM, Frost AJ, Lee CW, Papadimitriou JM, Dawkins HJS (1998): Development of PCR assays for species- and type-specific identification of Pasteurella multocida isolates. J Clin Microbiol 36: 1096–1100.
Townsend KM, Boyce JD, Chung JY, Frost AJ, Adler B (2001): Genetic organization of Pasteurella multocida cap Loci and development of a multiplex capsular PCR typing system. J Clin Microbiol 39: 924–929.
TSK Sachsen (2018): Jahresbericht 2018. Sächsische Tierseuchenkasse. Anstalt des öffentlichen Rechts. https://www.tsk-sachsen.de/documents/Jahresberichte/2019/Jahresbericht_….
Ujvári B, Szeredi L, Pertl L, Tóth G, Erdélyi K, Jánosi S, Molnár T, Magyar T (2015): First detection of Pasteurella multocida type B:2 in Hungary associated with systemic pasteurellosis in backyard pigs. Acta Vet Hung 63: 141–156.
Verma S, Sharma M, Katoch S, Verma L, Kumar S, Dogra V, Chahota R, Dhar P, Singh G (2013): Profiling of virulence associated genes of Pasteurella multocida isolated from cattle. Vet Res Commun 37: 83–89.
Völker I, Kehler W, Hewicker-Trautwein M, Seehusen F, Verspohl J, Bilk S, Baumgärtner W (2014): Re-emergence of haemorrhagic septicaemia in ungulates in Lower-Saxony in Germany. Vet Rec 175: 460.
WAHIS (2019): World Animal Health Information Database (WAHIS Interface) – Version 1. World Organisation for Animal Health (OIE). https://www.oie.int/wahis_2/public/wahid.php/Countryinformation/Country….

Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF 10.23761439-0299-2020-8-Sting.pdf (0.18 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF 10.23761439-0299-2020-8-Sting-Tabelle1.pdf (0.12 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF 10.23761439-0299-2020-8-Sting-Tabelle2.pdf (0.12 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF 10.23761439-0299-2020-8-Sting-Tabelle3.pdf (0.12 MB) herunterladen möchten




{kind=link}
{kind=link}