
Infektiöse Bronchitis: 80 Jahre Bekämpfungsanstrengungen zur Kontrolle einer Coronavirus-Infektion beim Geflügel
Berliner und Münchener Tierärztliche Wochenschrift 134, 1-8
DOI: 10.2376/1439-0299-2020-27
© Schlütersche Fachmedien GmbH. 2021
Publiziert: 01/2021
Summary
Infectious bronchitis (IB) is the most important Coronavirus infection in poultry. The IB virus (IBV) spreads rapidly in the affected flocks, and although airborne-spread is the principle route of transmission, contact also plays an important epidemiological role. This Gammacoronavirus infects susceptible chickens at all age groups. It affects not only the respiratory tract, but may also affect the urogenital and possibly to some extent the intestinal tract, depending on the strain characteristics. Besides strict biosecurity measures, vaccination is the most important strategy to control clinical disease. Vaccines, mainly live attenuated vaccines, have been used for almost 80 years. The high mutation and recombination rates and the frequent emergence of new virus variants in association with poor cross-protection, provide a continuous pressure to develop new vaccine candidates and strategies. Therefore, IB is a disease chickens will continue to be challenged with, and eradiation seems to be unrealistic at this point. The availability of advanced molecular techniques is currently opening up new opportunities to develop new generation vaccines and respond to newly emerging strains more quickly compared to traditional approaches. It is important that these new vaccine candidates stimulate the necessary immune mechanisms to provide protection at the main virus entrance site, the respiratory tract. Therefore, they should induce mucosal immunity including humoral as well as cell mediated reactions. The practicability for mass vaccination has to be ensured to be able to use these new vaccines in poultry production.
Zusammenfassung
Die Infektiöse Bronchitis (IB) ist die bedeutsamste Coronavirus-Erkrankung beim Nutzgeflügel. Das Virus der Infektiösen Bronchitis (IBV) breitet sich schnell in den betroffenen Beständen aus. Die Übertragung erfolgt insbesondere luftgetragen, aber auch Kontaktinfektionen spielen eine wichtige epidemiologische Rolle. Dieses Gamma-Coronavirus infiziert empfängliche Hühner aller Altersstufen und führt nicht nur zu Atemwegssymptomen, sondern kann auch, je nach Viruseigenschaften, zur Erkrankung des Urogenitaltrakts und bei einigen Stämmen auch des Darmtrakts führen. Neben strikten Biosicherheitsmaßnahmen stellen Impfungen die wichtigste Kontrollmöglichkeit der IB dar. Seit nahezu 80 Jahren werden Impfstoffe, insbesondere Lebendimpfstoffe, zur Kontrolle der IB eingesetzt. Jedoch führen die hohe Mutationsrate mit den daraus resultierenden neu entstehenden Virusvarianten sowie die meist schlechte Kreuzprotektion zu kontinuierlichen Herausforderungen und der wiederkehrenden Notwendigkeit, neue Impfstoffe zu entwickeln. Deshalb ist davon auszugehen, dass auch in Zukunft IB weiterhin eine gesundheitliche Herausforderung für unser Nutzgeflügel darstellen wird und eine Eradikation zu diesem Zeitpunkt unrealistisch erscheint. Die Möglichkeiten, auch molekularbiologische Techniken bei der Herstellung von Impfstoffkandidaten einzusetzen, werden es in Zukunft erlauben, neue Generationen von Impfstoffen gegen IB effektiv und vor allem schneller als mit den herkömmlichen Methoden zu entwickeln. Jedoch muss sichergestellt werden, dass diese Impfstoffe auch die notwendigen protektiven Immunmechanismen stimulieren, was bei IB insbesondere die Stimulation der mukosalen Immunität an der Viruseintrittspforte, dem Respirationstrakt, unter Einschluss der humoralen und zellvermittelten Parameter bedeutet. Weiterhin ist es für das Nutzgeflügel wichtig, dass auch diese neuen Impfstoffe für die Applikation bei großen Beständen geeignet sind.
Introduction
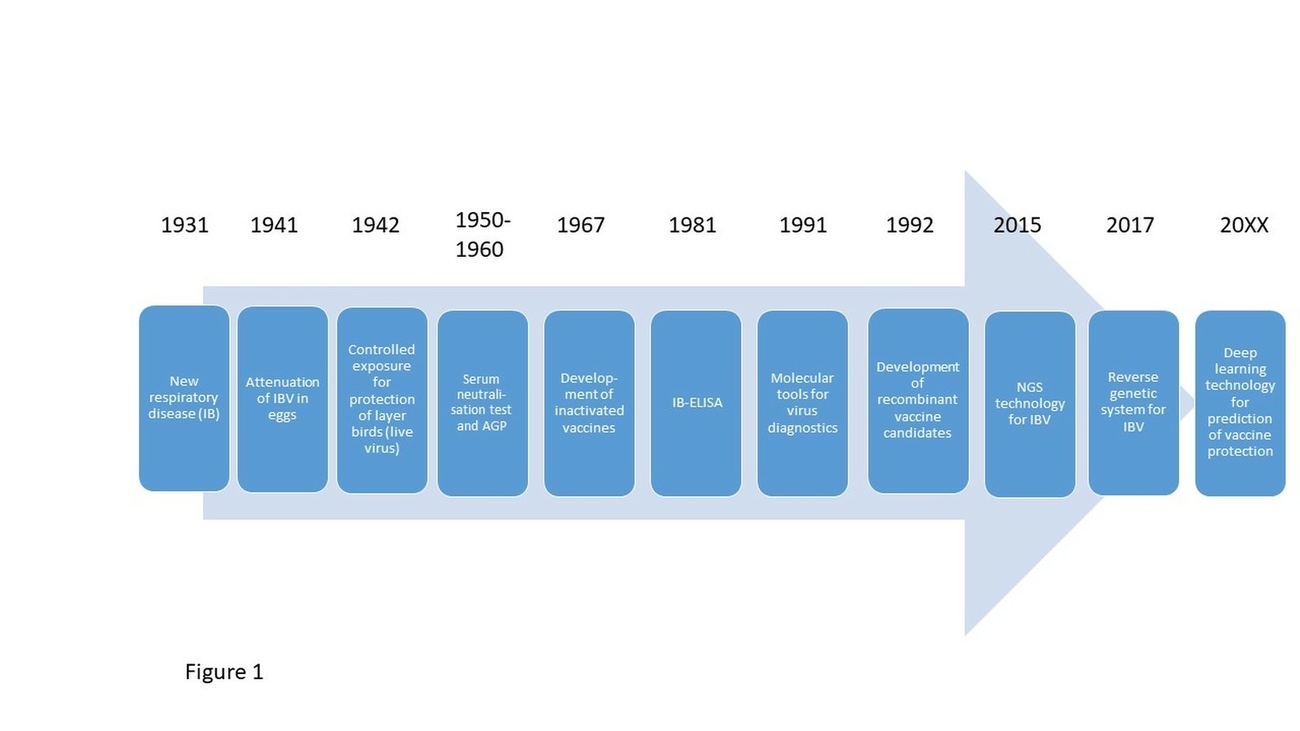
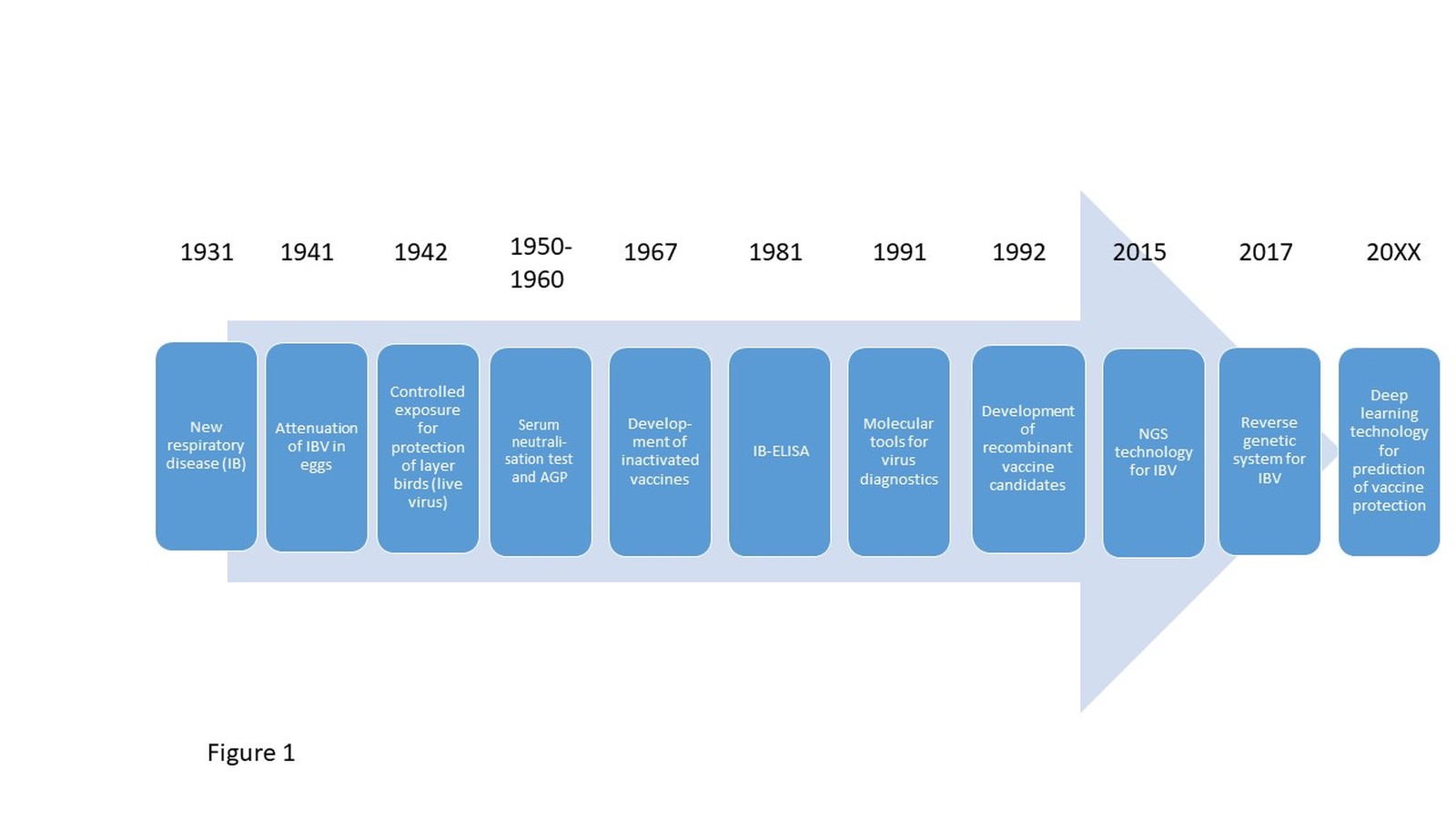
Infectious bronchitis (IB) is one of the most important respiratory disease in chickens. The destruction of the mucosa after infection with infectious bronchitis virus (IBV) allows the invasion of secondary pathogens, exacerbating the disease. The causing agent is a member of the species avian Coronavirus, which belongs to the genus Gammacoronavirus and family of Coronaviridae. IB was first observed in 1930 in the USA (Schalk and Hawn 1931) (Fig. 1). Today the virus is detected in almost all regions of the world (Bande et al. 2017). Some variants appear regionally restricted and only for a short time period, while others persist and spread (Legnardi et al. 2020). IBV and other avian Coronaviruses similar to IBV including Coronaviruses of pheasants, turkeys, ducks, goose and pigeon may affect multiple bird species, including wild birds, which may contribute to the spread of the virus (Cavanagh 2005). The Turkey Coronavirus (TCoV) may also lead to significant economic losses in turkeys, but IBV is by far the most important avian Gammacoronavirus worldwide (Jackwood and de Wit 2020). In this short review, we will focus on IBV and its main host species, the chicken. In the context of the newly emerging COVID-19 we would like to present lessons to be learned from many years of IBV research, featuring a Coronavirus of chickens also causing respiratory disease. We will specifically emphasize on preventive measures, their limitations and challenges.
The genome of IBV encodes for four structural proteins: spike (S), envelope (E), membrane (M), nucleocapsid (N), and several non-structural proteins (Jackwood and de Wit 2020). The spike protein is of specific interest for the virus-host interaction, as it forms projections on the virions. The S protein is post-translationally cleaved. The S1 unit contains hypervariable regions that play a role in the host receptor attachment, while the S2 unit is involved in membrane fusion and viral entry into the host cells. The S1 unit also provides conformational dependent virus-neutralizing epitopes (Cavanagh et al. 1998). IBV are traditionally grouped into serotypes by neutralization assays (Cook et al. 1999), and newly emerging serotypes are referred to as “variants”. As also known for other coronaviruses, IBV shows high recombination rates, most frequently observed in the nonstructural proteins 2, 3 and 16 as well as in the spike glycoprotein (Thor et al. 2011). This is mediated by a copy-choice mechanism (Legnardi et al. 2020). Typical for IBV is a high mutation rate resulting from poor viral polymerase proofreading, again especially in the S1 spike protein region. This explains the frequent development of new serotypes or strains, eventually capable of crossing species barriers (Jackwood et al. 2010). An average rate of 1.2 x 10–3 substitutions/site/year may occur in Coronaviruses (Hanada et al. 2004). Therefore, today sequencing of the S1 protein is frequently done to determine the genotype of the circulating IBVs. An amino acid similarity of greater than 90% is often associated with serological relatedness, but exceptions exist (Jackwood and de Wit 2020). Similarity based on the S1 protein sequence suggests cross-protection (de Wit et al. 2011). Based on the S1 gene phylogeny, Valastro et al. (2016) recently defined six genetic IBV types and 32 lineages worldwide, which can display different geographical distribution. The persistence and spread of new variants is suggested to be driven by environmental factors (Holmes 2009), including the microenvironment in different host tissues (Gallardo et al. 2010; Tab. 1). The study by Xu et al. (2018) demonstrated nicely the fast evolution of IBV in the field and the emergence of new sublineages, recombinants and variants in addition to the appearance of vaccine-like viruses within a short time period of only two years.
IB – a disease with different faces
Top Job:
IBV is very contagious. While the airborne route is epidemiologically most important, contact infections also occur. The infection spreads quickly within a group of chickens. Even vaccine strains may have an estimated reproduction ratio (R) of over 1, increasing the risk of vaccine virus spread between farms and the establishment of an endemic situation in a poultry population (Matthijs et al. 2008). There are only few indication of vertical transmission. The virus may be present on the egg-shell through shedding from the intestinal or urinary tract (Cook 1971, Cook et al. 2012, Jackwood and de Wit 2013). The highest virus concentration can be found in the trachea during the acute phase of infection. During the chronic stage, IBV may be detected in the intestinal tract, specifically in the caecal tonsils (de Wit 2000). Possible long-term excretion supports transmission between flocks by farm workers, contaminated litter, and other carrier material (Jackwood and de Wit 2020).
Infections with IBV may lead to different clinical pictures in susceptible chickens. The virus stain influences the type of symptoms. Furthermore, the age and sex, as well as the immune status of the host may have an impact on the disease development. Most IBV strains cause clinical infections of the upper respiratory tract in chickens of any age. IBV infection is often followed by secondary bacterial and viral infections complicating the course of the disease (Dwars et al. 2009). The morbidity is usually high to very high. It reaches up to 100% within only a few days, with variable but mostly low mortality rates after mono-infection. Urogenital manifestations may also be observed, especially in very young birds without maternally derived antibodies. This organ manifestation leads to considerable mortality and/or reduction in egg production in laying hens (Raj and Jones 1997). The so called “false layers” can be found in flocks starting the onset of lay, and are a result of neonatal infection of chickens with a serotype, against which no maternally derived immunity existed at the time of infection. Atresia of the oviduct is a result of the early IB infection, and becomes clinically apparent after sexual maturation of the pullet: the accumulation of albumen and retained ova lead to a massive swelling of the lower abdomen that manifests in a penguin-like posture of affected hens (Parent et al. 2020).
The spike protein determines the tissue tropism of IBV (Wickramasinghe et al. 2014). While the cellular receptors for IBV are still not fully elucidated, functional and structural data on the receptor binding domain of the spike protein are available (Shang et al. 2018). In vitro binding assays using cell-expressed IBV spike protein were established to study the affinity of IBV spikes to different tissues. The description of an enterotropic IBV strain by Saiada et al. (2020) underpins that even genetically closely related IBV strains may have diverging tissue tropism, calling for additional research on IBV structural biology. In the same manner, the estimation of the virulence of a given IBV strain is not possible solely based on serotyping and genotyping results. Pathotyping requires experimental in vitro and in vivo assessment using susceptible chickens and cell or organ culture systems, respectively (de Wit et al. 2017, Raj and Jones 1996).
Routine diagnostics
Rapid nucleic acid detection methods, mostly reverse transcriptase PCR (RT-PCR), are routinely used these days, to monitor chicken flocks and confirm the clinical suspicion of IB (Meir et al. 2010). Poultry veterinary services, which are offered on a regular basis to commercial farms, also provide access to these rapid diagnostic technologies.
The molecular techniques allow investigating infectious disease outbreaks timely and efficiently, which is the foundation for the application of efficient disease control measures and is important to prevent additional health problems with economic losses associated with secondary infections. The factors leading to a fast evolution of modern molecular diagnostic tools in poultry production and their field application have been manifold. Oropharyngeal swabs, which are suitable for the diagnosis of respiratory infections, are easy to collect, and can be pooled to reduce the cost of analysis (Zwaagstra et al. 1992). Molecular reagents, test kits, and laboratory hardware are now cost-effective and easy to handle. Samples can be stored and transported conveniently using FTA-cards, even allowing for safe and convenient international shipment (Manswr et al. 2018). If virus propagation is needed to be done (for example for the preparation of autogenous vaccines), cold chains have to be maintained for sample transport, as the virus is not very stable at higher temperatures (Jackwood and de Wit 2020).
A typical diagnostic approach to detect IB would start with the detection of the RNA of the causative agent using a species-specific RT-PCR, or a multiplex PCR approach focusing on IBV genotypes of particular relevance for the respective geographic region. Further differentiation of positive findings is performed by sequencing (Jiang et al. 2017). Next generation sequencing (NGS) has some advantages over classical protocols, as it is an untargeted method and thus not limited to agents expected to be present in the sample. Furthermore, NGS can pick up virus subpopulations that remain undetected when classical sequencing methods are used (Tang et al. 2016). However, NGS has not become a routine method yet in veterinary diagnostics.
While nucleic-acid detection based diagnostics allow for a quick overview on the prevalence of IBV in chicken flocks, virus isolation remains the key for a thorough understanding of IBV field strains, and is necessary to determine their potential role in emerging diseases through pathotyping. As IBV field isolates cannot be cultured on standard laboratory cell lines, the procedure is tedious and expensive. A current example for a full-scale examination of emerging IBV in a specific geographic is given by Ren et al. (2020). The authors studied field isolates from relevant cases in vivo and in vitro, and identified virus strains that differ in virulence and antigenicity while appearing similar genetically.
Control strategies
General biosecurity measures including proper cleaning and disinfection are important to reduce the IBV field pressure. As IBV is an enveloped virus, most common disinfectants, which are used in poultry houses, inactivate the virus (Jackwood and de Wit 2020). Due to the fact of IBV being easily transmissible and widely distributed, in many countries immunization of commercial poultry is necessary to prevent clinical disease and production impairment. Depending on the vaccine type, immunization takes place by different routes: eye drop, course spray, drinking water, and aerosol (De Wit et al. 2010). Spray vaccination is the most common way, and may already take place in the hatchery. An early IB-vaccination at one day of age seems to induce efficient immune responses. This method is easy to standardize in the hatchery, reaches a high coverage rate and induces very early immune responses helping to overcome the persistent field pressure (Jackwood et al. 2020, Legnardi et al. 2020). On the other hand, recent studies suggested that birds at seven days of age mount a better local humoral and cell-mediated immunity after vaccination with some IB-vaccine strains compared to one-day old chicks (Zegpi et al. 2020a). IB-vaccination also may be combined with vaccines against other pathogens.
Maternal antibodies hardly interfere with challenge virus infection (Hofstad and Kenzy 1950) or vaccine take. This supports the fact that circulating antibodies do not play a major role in IB control. While the absence of detectable circulating antibodies after IBV vaccination was not associated with lack of protection (Chubb 1974), high levels of circulating antibodies did not always provide sufficient protection against challenge (Pensaert and Lambrechts 1994).
Overall, local innate, cellular, and humoral responses are most important in IB-protection (Chhabra et al. 2015a, b). Besides the role of local humoral immunity for virus control, specifically IgA in lacrimal fluid and trachea, also cell-mediated immune mechanisms seem to contribute to the initial elimination of the virus from the lung and kidney. Experimental studies provided evidence that cytotoxic T lymphocytes protect the respiratory tract from acute infection (Collisson et al. 2000, Seo et al. 2000).
IBV-induced innate immune responses vary significantly between chicken genotypes, which may associate with the Major Histocompatibility Complex. While the susceptible genotype mounted a detrimental local inflammatory response in the trachea within the early hours post inoculation, the more resistant birds showed a lower baseline cytokine expression and subsequently tissue lesions were less pronounced after IBV-challenge (da Silva et al. 2020). This difference in innate immune response did not seem to be related to the magnitude and quality of the adaptive immune response (da Silva et al. 2020).
The experimental use of a live IBV vaccine demonstrated a significant reduction of the challenge virus transmission rate from an estimated R > 19 to an estimated R = 0.69 (de Wit et al. 1998). Similar observations were also made by Matthijs et al. (2008), confirming the reduction of the R value below 1 in vaccinated birds challenged with a more virulent virus of the same serotype. Besides classical attenuated live also inactivated vaccines (usually oil-adjuvanted) are licensed on the international market. The first IBV live “vaccination approach” was reported in the 1940th (van Roeckel et al. 1942) (Fig. 1). Live vaccines are normally used in meat type birds, and to prime layer as well as breeder pullets. Booster vaccination in breeder and layer flocks includes the use of inactivated vaccines that are administered before lay. These inactivated vaccines induce significant humoral immunity, which in this case protects against IBV-induced egg-quality changes and kidney lesions. Unfortunately, inactivated vaccines do not induce protection of the respiratory tract, and furthermore, the overall protection at the local epithelial target tissue sites is only of short duration (Cook et al. 1986). Therefore, chickens, especially layers and breeders in regions with a high infectious pressure, are continuously boostered with live vaccines, to maintain local protection in the respiratory tract, which is the site of virus entry.
Already early in the 1950th it was clear that various serotypes and antigen variants circulate and continue to emerge in the field, and vaccines derived from other strains with different antigenic properties may only provide poor or no protection (de Wit et al. 2011, Jordan 2017, Jungherr et al. 1956). It was deemed unrealistic and inefficient to develop homologous vaccines against a plethora of IBV serotypes. Systematic cross protection studies in animals demonstrated the efficacy of certain vaccines and combinations thereof against challenge infection with IBV variants, leading to the concept of “protectotypes” and “protectotyping” (Bande et al. 2015, Cook et al. 1999). To date, protectoyping is used to identify the “best fit” vaccination program for a given IBV field situation based on the existing vaccines, rather than directly committing to the development of a new product (de Wit et al. 2017).
The more diversified and heterogeneous immune response spectrum may have the advantage of interfering with the emergence of specific immunity escape mutants (Franzo et al. 2019). On the other hand though, this approach may pave the way for recombinations. Birds may be co-infected with different IB viruses at the same time including recombinants of vaccine-derived strains (Han et al. 2018).
At the moment there are about 12 inactivated vaccines licensed in Germany, which contain an IBV in combination with different other pathogens. The IBV component may consist of one IBV serotype or two antigenically different types (PEI 2020). In addition, about 15 live vaccines are on the market including at least one of the different IBV serotypes of importance in Europe such as Massachusetts type, 4/91, D276 or QX-like viruses (Bande et al. 2017).
Due to the high variability of field strains, licensed vaccines may not sufficiently protect against some newly emerging antigenic variants. As the development of new vaccines may take a few years from virus isolation up to the release of a licensed product, also autogenous vaccines are used. Normally these are inactivated vaccines, which reduce the risk of further spread of the newly emerging variant strains.
Advances in molecular biology allowed for the development of reverse genetic systems for IBV (van Beurden et al. 2017; Fig. 1). This led together with the identification of suitable vectors for recombinant vaccines and subunit vaccines, which are prepared on the basis of different spike protein ectodomains, to the development of new generation vaccines (Eldemery et al. 2017, Tarpey et al. 2007, Toro et al. 2014, Zegpi et al. 2020b). Also DNA vaccines encoding N or S1 genes have been tested, some of which showed better performance if cytokines or chemokines are encoded as well (Tang et al. 2008, Tian et al. 2008). Until now, none of these vaccine candidates are licensed, as they do not provide advantages over conventional live vaccines. Most of them show insufficient cross protection (Keep et al. 2020) and/or require individual bird vaccination, which is not feasible in the field.
In addition to the properties of the vaccine, vaccination success largely depends on the quality of vaccine administration. de Wit et al. (2010) identified factors in association with the vaccine administration process that may influence the quality of the immune response after vaccination, including the genotype of the birds, age and time intervals between booster vaccinations. Evidence exists that good vaccination practice leads to better immunization results (de Wit et al. 2010). Co-infections with immunosuppressive pathogens (van Ginkel et al. 2008) or vaccination with additional vaccines against different pathogens may interfere with an appropriate immune response. On the other hand the co-application of a Newcastle disease virus, an Avian Metapneumovirus and two live IBV vaccines to day-old commercial broiler chickens did not affect IBV-immunity (Ball et al. 2019).
Success story of prevention?
The first vaccinations against IBV were carried out as early as 1942 (Fig. 1), and are now common practice around the world for all kinds of chicken production. The emergence of new IBV serotypes regularly calls for the development of updated live vaccines, and there is no evidence for the eradication of IB from the field. Franzo et al. (2019) recently demonstrated and confirmed that the introduction of an IB vaccine belonging to a new serotype puts evolutionary pressure on the field virus population (Tab. 1). The authors could even identify epitopes in the IB spike protein that are particularly prone to changes when a homologous vaccine is present. In spite of the good clinical efficacy of IB vaccination against a homologous challenge, available vaccines can only reduce but not completely block viral shedding. Sterile immunity is not achievable, although vaccination may reduce viral shedding of homologous viruses by more than 1 log10 of excreted titer (Legnardi et al. 2020). Studies in the US furthermore indicate that mass vaccination additionally contributes to further spread of vaccine-like viruses, as new subpopulations may emerge (Toro et al. 2012b) (Tab. 1). Often insufficient vaccination technology leads to an initial infection of less than 60% of the vaccinated chickens. But the vaccine virus was shown to spread quickly to the other birds of the flock increasing the chance for reversion to more virulence when bird-to-bird transmission takes place (Callison et al. 2006). In some field studies most positive IBV findings were related to vaccines, and only 17% were attributed to IBV field strains (Legnardi et al. 2019). Therefore, the success of an IB-vaccination strategy does not only depend on the vaccine virus or combination of vaccine viruses and proper vaccination technology, also monitoring and evaluation of the vaccination impact in the specific region in the face of the circulating IBV strains is important for the ongoing improvement of prophylactic strategies (Franzo et al. 2016).
Another factor limiting the success of current IB vaccination in the field is the circulation of many different serotypes in a certain region. Cross-protection between different serotypes is generally limited to low. Therefore, we can conclude from all the efforts to control IBV for over 80 years, that there is still much to be learned as we did not succeed in eradicating the virus yet. At the moment live vaccines seem to work best to control the clinical respiratory disease and reduce shedding as they induce the necessary local immune reactions. However, we know that this local immunity is of short duration making frequent booster vaccination necessary in the field. New diagnostic strategies allow rapid identification of IBV and most importantly the identification of new genotypes, variants and recombinants by sequencing approaches. This in combination with the use of reverse genetic systems will allow the development of new generation vaccines, eventually targeted vaccines as future candidates for the induction of protection against newly evolving IBVs.
Conflict of interest
The authors hereby declare that they have no proprietary, professional or other personal interests in any product, service and/or company that could have influenced the contents or opinions expressed in this publication.
Authors contribution
Both authors contributed to the manuscript evenly.
Address for correspondence
Prof. Dr. Silke Rautenschlein
Klinik für Geflügel
Stiftung Tierärztliche Hochschule Hannover
Bünteweg 9
30559 Hannover
Silke.Rautenschlein@tiho-hannover.de
Literature
Ball C, Forrester A, Herrmann A, Lemiere S, Ganapathy K (2019): Comparative protective immunity provided by live vaccines of Newcastle disease virus or avian metapneumovirus when co-administered alongside classical and variant strains of infectious bronchitis virus in day-old broiler chicks. Vaccine 37: 7566–7575.
Bande F, Arshad SS, Bejo MH, Moeini H, Omar AR (2015): Progress and challenges toward the development of vaccines against avian infectious bronchitis. J Immunol Res 2015: 424860.
Bande F, Arshad SS, Omar AR, Hair-Bejo M, Mahmuda A, Nair V (2017): Global distributions and strain diversity of avian infectious bronchitis virus: a review. Anim Health Res Rev 18: 70–83.
Callison SA, Hilt DA, Boynton TO, Sample BF, Robison R, Swayne DE, Jackwood MW (2006): Development and evaluation of a real-time Taqman RT-PCR assay for the detection of infectious bronchitis virus from infected chickens. J Virol Methods 138: 60–65.
Cavanagh D (2005): Coronaviruses in poultry and other birds. Avian Pathol 34: 439–448.
Cavanagh D (2007): Coronavirus avian infectious bronchitis virus. Vet Res 38: 281–297.
Cavanagh D, Mawditt K, Adzhar A, Gough RE, Picault JP, Naylor CJ, Haydon D, Shaw K, Britton P (1998): Does IBV change slowly despite the capacity of the spike protein to vary greatly? Adv Exp Med Biol 440: 729–734.
Chhabra R, Chantrey J, Ganapathy K (2015a): Immune Responses to Virulent and Vaccine Strains of Infectious Bronchitis Viruses in Chickens. Viral Immunol 28: 478–488.
Chhabra R, Forrester A, Lemiere S, Awad F, Chantrey J, Ganapathy K (2015b): Mucosal, Cellular, and Humoral Immune Responses Induced by Different Live Infectious Bronchitis Virus Vaccination Regimes and Protection Conferred against Infectious Bronchitis Virus Q1 Strain. Clin Vaccine Immunol 22: 1050–1059.
Chubb RC (1974): The effect of the suppression of circulating antibody on resistance to the Australian avian infectious bronchitis virus. Res Vet Sci 17: 169–173.
Collisson EW, Pei J, Dzielawa J, Seo SH (2000): Cytotoxic T lymphocytes are critical in the control of infectious bronchitis virus in poultry. Dev Comp Immunol 24: 187–200.
Cook JK 1971: Recovery of infectious bronchitis virus from eggs and chicks produced by experimentally inoculated hens. J Comp Pathol 81: 203–211.
Cook JK, Smith HW, Huggins MB (1986): Infectious bronchitis immunity: its study in chickens experimentally infected with mixtures of infectious bronchitis virus and Escherichia coli. J Gen Virol 67(Pt 7): 1427–1434.
Cook JK, Orbell SJ, Woods MA, Huggins MB (1999): Breadth of protection of the respiratory tract provided by different live – attenuated infectious bronchitis vaccines against challenge with infectious bronchitis viruses of heterologous serotypes. Avian Pathol 28: 477–485.
Cook JK, Jackwood M, Jones RC (2012): The long view:
40 years of infectious bronchitis research. Avian Pathol 41: 239–250.
da Silva AP, Schat KA, Gallardo RA (2020): Cytokine Responses in Tracheas from Major Histocompatibility Complex Congenic Chicken Lines with Distinct Susceptibilities to Infectious Bronchitis Virus. Avian Dis 64: 36–45.
de Wit JJ (2000): Detection of infectious bronchitis virus. Avian Pathol 29: 71–93.
de Wit JJ, de Jong, MC, Pijpers A, Verheijden, JH (1998): Transmission of infectious bronchitis virus within vaccinated
and unvaccinated groups of chickens. Avian Pathol 27: 464–471.
de Wit JJ, Swart WA, Fabri TH (2010): Efficacy of infectious bronchitis virus vaccinations in the field: association between the alpha-IBV IgM response, protection and vaccine application parameters. Avian Pathol 39: 123–131.
de Wit JJ, Cook JK, van der Heijden HM (2011): Infectious bronchitis virus variants: a review of the history, current situation and control measures. Avian Pathol 40: 223–235.
de Wit JJ, Dijkman R, Clavo J, Gomzales A, Hidalgo H (2017): Variability in biological beahviour, pathogenicity, protectotype and induction of virus neutralizing antibodies by different vaccination programmes to infectious bronchitis virus genotype Q1 strains from Chile. Avian Pathol 46: 666–675.
Dwars RM, Matthijs MG, Daemen AJ, van Eck JH, Vervelde L, Landman WJ (2009): Progression of lesions in the respiratory tract of broilers after single infection with Escherichia coli
compared to superinfection with E. coli after infection with infectious bronchitis virus. Vet Immunol Immunopathol 127: 65–76.
Eldemery F, Joiner KS, Toro H, van Santen VL (2017): Protection against infectious bronchitis virus by spike ectodomain subunit vaccine. Vaccine 35: 5864–5871.
Franzo G, Tucciarone CM, Blanco A, Nofrarías M, Biarnés M, Cortey M, Majó N, Catelli E, Cecchinato M (2016): Effect of different vaccination strategies on IBV QX population dynamics and clinical outbreaks. Vaccine 34: 5670–5676.
Franzo G, Massi P, Tucciarone CM, Barbieri I, Tosi G, Fiorentini L, Ciccozzi M, Lavazza A, Cecchinato M, Moreno A (2017): Think globally, act locally: Phylodynamic reconstruction of infectious bronchitis virus (IBV) QX genotype (GI-19 lineage) reveals different population dynamics and spreading patterns when evaluated on different epidemiological scales. PLoS One 12: e0184401.
Franzo G, Legnardi M, Tucciarone CM, Drigo M, Martini M, Cecchinato M (2019): Evolution of infectious bronchitis virus in the field after homologous vaccination introduction. Vet Res 50: 92.
Gallardo RA, van Santen VL, Toro H (2010): Host intraspatial selection of infectious bronchitis virus populations. Avian Dis 54: 807–813.
Guzman M, Hidalgo H (2020): Live Attenuated Infectious Bronchitis Virus Vaccines in Poultry: Modifying Local Viral Populations Dynamics. Animals (Basel) 10: 2058.
Han Z, Gao M, Chen Y, Zhao W, Sun J, Zhao Y, Liu S (2018): Genetics, antigenicity and virulence properties of three infectious bronchitis viruses isolated from a single tracheal sample in a chicken with respiratory problems. Virus Res 257: 82–93.
Hanada K, Suzuki Y, Gojobori T (2004): A large variation in the rates of synonymous substitution for RNA viruses and its relationship to a diversity of viral infection and transmission modes. Mol Biol Evol 21: 1074–1080.
Hassan KE, Ali A, Shany SAS, El-Kady MF (2017): Experimental co-infection of infectious bronchitis and low pathogenic avian influenza H9N2 viruses in commercial broiler chickens. Res Vet Sci 115: 356–362.
Hofstad MS, Kenzy SG (1950): Susceptibility of chicks hatched from recovered hens to infectious bronchitis. Cornell Vet 40: 87–89.
Holmes EC (2009): Evolution and emergence of RNA viruses. Oxford University Press, New York.
Jackwood MW, de Wit JJ (2013): Infectious bronchitis. In: Swayne DE (ed.), Diseases of Poultry. 13th ed. Wiley Blackwell, Hoboken, NJ, USA, 139–159.
Jackwood MW, de Wit JJ (2020): Infectious bronchitis. In: Swayne DE, Boulianne M, McDougald LR, Nair V, and Suarez DL (eds.), Diseases of Poultry. Wiley Blackwell, Hoboken, NJ, USA, 167–188.
Jackwood MW, Hilt DA, McCall AW, Polizzi CN, McKinley ET, Williams SM (2009): Infectious bronchitis virus field vaccination coverage and persistence of Arkansas-type viruses in commercial broilers. Avian Dis 53: 175–183.
Jackwood MW, Boynton TO, Hilt DA, McKinley ET, Kissinger JC, Paterson AH, Robertson J, Lemke C, McCall AW, Williams SM, Jackwood JW, Byrd LA (2010): Emergence of a group 3 coronavirus through recombination. Virology 398: 98–108.
Jackwood MW, Hall D, Handel A (2012): Molecular evolution and emergence of avian gammacoronaviruses. Infect Genet Evol 122: 1305–1311.
Jackwood MW, Clark R, Cheng S, Jordan BJ (2020): Protection following simultaneous vaccination with three or four different attenuated live vaccine types against infectious bronchitis virus. Avian Pathol 49: 335–341.
Jiang L, Zhao W, Han Z, Chen Y, Zhao Y, Sun J, Li H, Shao Y, Liu L, Liu S (2017): Genome characterization, antigenicity and pathogenicity of a novel infectious bronchitis virus type isolated from south China. Infect Genet Evol 54: 437–446.
Jordan B (2017): Vaccination against infectious bronchitis virus: A continuous challenge. Vet Microbiol 206 137–143.
Jungherr EL, Chomiak TW, Luginbuhl RE (1956): Immunological differences in strains of infectious bronchitis virus. Chicago, IL, USA, 203–209.
Keep S, Sives S, Stevenson-Leggett P, Britton P, Vervelde L, Bickerton E (2020): Limited Cross-Protection against Infectious Bronchitis Provided by Recombinant Infectious Bronchitis Viruses Expressing Heterologous Spike Glycoproteins. Vaccines (Basel) 8(2): 330.
Legnardi M, Franzo G, Koutoulis KC, Wiśniewski M, Catelli E, Tucciarone CM, Cecchinato M (2019): Vaccine or field strains: the jigsaw pattern of infectious bronchitis virus molecular epidemiology in Poland. Poult Sci 98: 6388–6392.
Legnardi M, Tucciarone CM, Franzo G, Cecchinato M (2020): Infectious Bronchitis Virus Evolution, Diagnosis and Control. Vet Sci 7(2): 79.
Manswr B, Ball C, Forrester A, Chantrey J, Ganapathy K (2018): Evaluation of full S1 gene sequencing of classical and variant infectious bronchitis viruses extracted from allantoic fluid and FTA cards. Avian Pathol 47: 418–426.
Matthijs MGR, Bouma A, Velkers FC, van Eck JHH, Stegeman JA (2008): Transmissibility of infectious bronchitis virus H120 vaccine strain among broilers under experimental conditions. Avian Dis 52: 461–466.
Meir R, Maharat O, Farnushi Y, d Simanov L (2010): Development of a real-time TaqMan RT-PCR assay for the detection of infectious bronchitis virus in chickens, and comparison of RT-PCR and virus isolation. J Virol Methods 163: 190–194.
Parent E, Gagnon-Francoeur A, Lanthier B, Hebert G, Buczinski S, d Boulianne M (2020): Diagnostic Accuracy of Ultrasonography to Detect False Layers in a Commercial Laying Flock Infected by an Infectious Bronchitis Virus Delmarva Genotype Causing Cystic Oviducts. Avian Dis 64: 149–156.
PEI (2020): https://www.pei.de/DE/arzneimittel/tierarzneimittel/gefluegel/gefluegel–node.html?cms_gts=11866844_list%253Dheader_text_sort%252Basc&cms_gtp=11866844_list%253D6; website visit: 04.08.2020.
Pensaert M, Lambrechts C (1994): Vaccination of chickens against a Belgian nephropathogenic strain of infectious bronchitis virus B1648 using attenuated homologous and heterologous strains. Avian Pathol 23: 631–641.
Raj GD, Jones RC (1996): An in vitro comparison of the virulence of seven strains of infectious bronchitis virus using tracheal and oviduct organ cultures. Avian Pathol 25: 649–662.
Raj GD, Jones RC (1997): Infectious bronchitis virus: Immunopathogenesis of infection in the chicken. Avian Pathol 26: 677–706.
Ren M, Zhang L, Hou Y, Zhao Y, Han Z, Sun J, Liu S (2020): Genetic, Antigenic, and Pathogenic Characteristics of Infectious Bronchitis Virus GI-7/TW-II in China. Avian Dis 64: 183–196.
Saiada F, Eldemery F, Zegpi RA, Gulley SL, Mishra A, van Santen VL, Toro H (2019): Early Vaccination of Chickens Induces Suboptimal Immunity Against Infectious Bronchitis Virus. Avian Dis 63: 38–47.
Saiada F, Gallardo RA, Shivaprasad HL, Corsiglia C, Van Santen VL (2020): Intestinal Tropism of an Infectious Bronchitis Virus Isolate Not Explained by Spike Protein Binding Specificity. Avian Dis 64: 23–35.
Schalk AF, Hawn MC (1931): An apparent new respiratory disease of baby chicks. J Am Vet Med Assoc 78: 413–422.
Seo SH, Pei J, Briles WE, Dzielawa J, Collisson EW (2000): Adoptive transfer of infectious bronchitis virus primed alphabeta T cells bearing CD8 antigen protects chicks from acute infection. Virology 269: 183–189.
Shang J, Zheng Y, Yang Y, Liu C, Geng Q, Luo C, Zhang W, Li F (2018): Cryo-EM structure of infectious bronchitis coronavirus spike protein reveals structural and functional evolution of coronavirus spike proteins. PLoS Pathog 14: e1007009.
Tang M, Wang H, Zhou S, Tian G (2008): Enhancement of the immunogenicity of an infectious bronchitis virus DNA vaccine by a bicistronic plasmid encoding nucleocapsid protein and interleukin–2. J Virol Methods 149: 42–48.
Tang Y, Lin L, Sebastian A, Lu H (2016): Detection and characterization of two co-infection variant strains of avian orthoreovirus (ARV) in young layer chickens using next-generation sequencing (NGS). Sci Rep 6: 24519.
Tarpey I, van Loon AA, de Haas N, Davis PJ, Orbell S, Cavanagh D, Britton P, Casais R, Sondermeijer P, Sundick R (2007): A recombinant turkey herpesvirus expressing chicken interleukin-2 increases the protection provided by in ovo vaccination with infectious bursal disease and infectious bronchitis virus. Vaccine 25: 8529–8535.
Thor SW, Hilt DA, Kissinger JC, Paterson AH, Jackwood MW (2011): Recombination in avian gamma-coronavirus infectious bronchitis virus. Viruses 3: 1777–1799.
Tian L, Wang HN, Lu D, Zhang YF, Wang T, Kang RM (2008): The immunoreactivity of a chimeric multi-epitope DNA vaccine against IBV in chickens. Biochem Biophys Res Commun 377: 221–225.
Toro H, van Santen VL, Jackwood MW (2012a): Genetic diversity and selection regulates evolution of infectious bronchitis virus. Avian Dis 56: 449–455.
Toro H, Pennington D, Gallardo RA, van Santen VL, van Ginkel FW, Zhang J, Joiner KS (2012b): Infectious bronchitis virus subpopulations in vaccinated chickens after challenge. Avian Dis 56: 501–508.
Toro H, Zhang JF, Gallardo RA, van Santen VL, van Ginkel FW, Joiner KS, Breedlove C (2014): S1 of distinct IBV population expressed from recombinant adenovirus confers protection against challenge. Avian Dis 58: 211–215.
Valastro V, Holmes EC, Britton P, Fusaro A, Jackwood MW, Cattoli G, Monne I (2016): S1 gene-based phylogeny of infectious bronchitis virus: An attempt to harmonize virus classification. Infect Genet Evol 39: 349–364.
van Beurden SJ, Berends AJ, Kramer-Kuhl A, Spekreijse D, Chenard G, Philipp HC, Mundt E, Rottier PJM, Verheije MH (2017): A reverse genetics system for avian coronavirus infectious bronchitis virus based on targeted RNA recombination. Virol J 14: 109.
van Ginkel FW, van Santen VL, Gulley SL, Toro H (2008): Infectious bronchitis virus in the chicken Harderian gland and lachrymal fluid: viral load, infectivity, immune cell responses, and effects of viral immunodeficiency. Avian Dis 52: 608–617.
van Roeckel H, Bullis KL, Flint OS, Clarke MK (1942): Poultry disease control service. Massachusetts Agricultural Experimental Station, MA Annual Report Bulletin, 99–103.
Wickramasinghe IN, van Beurden SJ, Weerts EA, Verheije MH (2014): The avian coronavirus spike protein. Virus Res 194: 37–48.
Xu L, Han Z, Jiang L, Sun J, Zhao Y, Liu S (2018): Genetic diversity of avian infectious bronchitis virus in China in recent years. Infect Genet Evol 66: 82–94.
Zegpi RA, Breedlove C, Gulley S, Toro H (2020a): Infectious Bronchitis Virus Immune Responses in the Harderian Gland upon Initial Vaccination. Avian Dis 64: 92–95.
Zegpi RA, He L, Yu Q, Joiner KS, van Santen VL, Toro H (2020b): Limited Protection Conferred by Recombinant Newcastle Disease Virus Expressing Infectious Bronchitis Spike Protein. Avian Dis 64: 53–59.
Zhao Y, Zhang H, Zhao J, Zhong Q, Jin J-H, Zhang G-Z (2016): Evolution of infectious bronchitis virus in China over the past two decades. J Gen Virol 97: 1566–1574.
Zwaagstra KA, van der Zeijst BA, Kusters JG (1992): Rapid detection and identification of avian infectious bronchitis virus. J Clin Microbiol 30: 79–84.

Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2020-27-Rautenschlein.pdf (0.15 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2020-27-Rautenschlein-Tabelle1.pdf (0.12 MB) herunterladen möchten



{kind=link}