

Disseminierte atypische Mykobakteriose durch Mycobacterium xenopi bei einem Weißkopfsaki (Pithecia pithecia)
Berliner und Münchener Tierärztliche Wochenschrift 135, 1–5
DOI: 10.2376/1439-0299-2021-24
© Schlütersche Fachmedien GmbH. 2022
Eingereicht: 4. November 2021
Akzeptiert: 20. Dezember 2021
Publiziert: 01/2022
Summary
A 15.5 year old male white-faced saki (Pithecia pithecia) was euthanized due to severe emaciation and an ultrasonographically suspected hepatic neoplasm. Postmortem examination revealed disseminated granulomatous inflammation affecting liver, pancreas, kidneys, heart, skin, and various lymph nodes. Mycobacteria were isolated from pulmonary lymph node and liver, identified as Mycobacterium (M.) xenopi a non-tuberculous mycobacterial species of human relevance especially in cases of underlying immunodeficiency. Subsequent characterization using ITS-sequencing and MLST-pattern analysis revealed that the isolate was identical with a Canadian strain of human origin indicating a high degree of conservation across countries and host species. This case represents to our knowledge the first report of a clinical M. xenopi infection in a host species not yet reported.
Zusammenfassung
Ein 15,5 Jahre alter männlicher Weißkopfsaki (Pithecia pithecia) aus einem Zoo wurde wegen hochgradiger Abmagerung und des sonografischen Verdachts auf eine Leberneoplasie euthanasiert. Bei der Sektion zeigten sich granulomatöse Herdveränderungen disseminiert in Leber, Pankreas, Nieren, Herz, Haut und verschiedenen Lymphknoten. Aus einem Lungenlymphknoten und der Leber wurde Mycobacterium (M.) xenopi isoliert. Dieser Erreger zählt zu den nichttuberkulösen Mykobakterien mit zoonotischer Relevanz, insbesondere bei immundefizienten Menschen. Bei der weitergehenden Erregerdifferenzierung mittels ITS-Sequenzierung und MLST-Pattern-Analyse erwies sich der Stamm als identisch mit einem humanen kanadischen Isolat. Dieses Ergebnis bestätigt bei dem Erreger einen hohen Konservierungsgrad über Ländergrenzen und Wirtspezies hinweg. Bei dem beschriebenen Fall handelt es sich unseres Wissens nach um die Erstbeschreibung einer klinisch manifesten M. xenopi-Infektion bei einem Weißkopfsaki.
Introduction
Generalized mycobacterial disease by non-tuberculous mycobacteria (NTM) is rarely reported in non-human primates. Especially New World monkeys are considered as highly resistant for mycobacterial infections (Mätz-Rensing and Kaup 2012). Sakis are medium-sized New World monkeys living in the Amazon region. There are only three reports on NTM infections in sakis. Mycobacterium intracellulare was isolated from a white-faced saki with multifocal granulomatous lymphadenitis kept in a zoo in Florida (Heard et al. 1997). In one of two black bearded sakis from a South Korean zoo with multifocal subcutaneous nodular lesions M. kansasii-infection was detected, whereas mycobacterial speciation failed in the other (Murai et al. 2012). Here, we report the disseminated M. xenopi-infection in a white-faced saki, kept in another German zoological garden and about the results of ITS-sequencing and MLST-pattern analysis of the isolated M. xenopi strain. This case represents a M. xenopi-infection in a non-human primate, a host species not yet reported.
Case description
Clinical signs and necropsy
Top Job:
A 15.5-year old male white-faced saki (Pithecia pithecia) with suprelorin® (Virbac AG, Switzerland) chip implantation since two years, showed reduced activity with retained appetite and had halved his body weight within a few month to 1.25 kg. The animal was born in captivity in France before moving to Zoo Dortmund (Germany) at an age of 4 years. Here the saki was kept in a group of white-faced sakis (Fig. 1) temporarily together with squirrel monkeys (Saimiri sciureus sciureus) and golden agoutis (Dasyprocta aguti) in an in/outdoor enclosure from 2004 until its death in 2016. In June 2016, the group of white-faced sakis consisted of 5 male (15.5, 4, 2.5, 1.5, and 1 years of age) and 3 female animals (14, 9, and 1 years of age). On the left cheek, there was a poorly healing skin ulceration of approximately 2.5 cm in diameter. A small lesion at this site had already been detected in September 2014, but was not further paid attention to because it did not seem to have any clinical relevance.
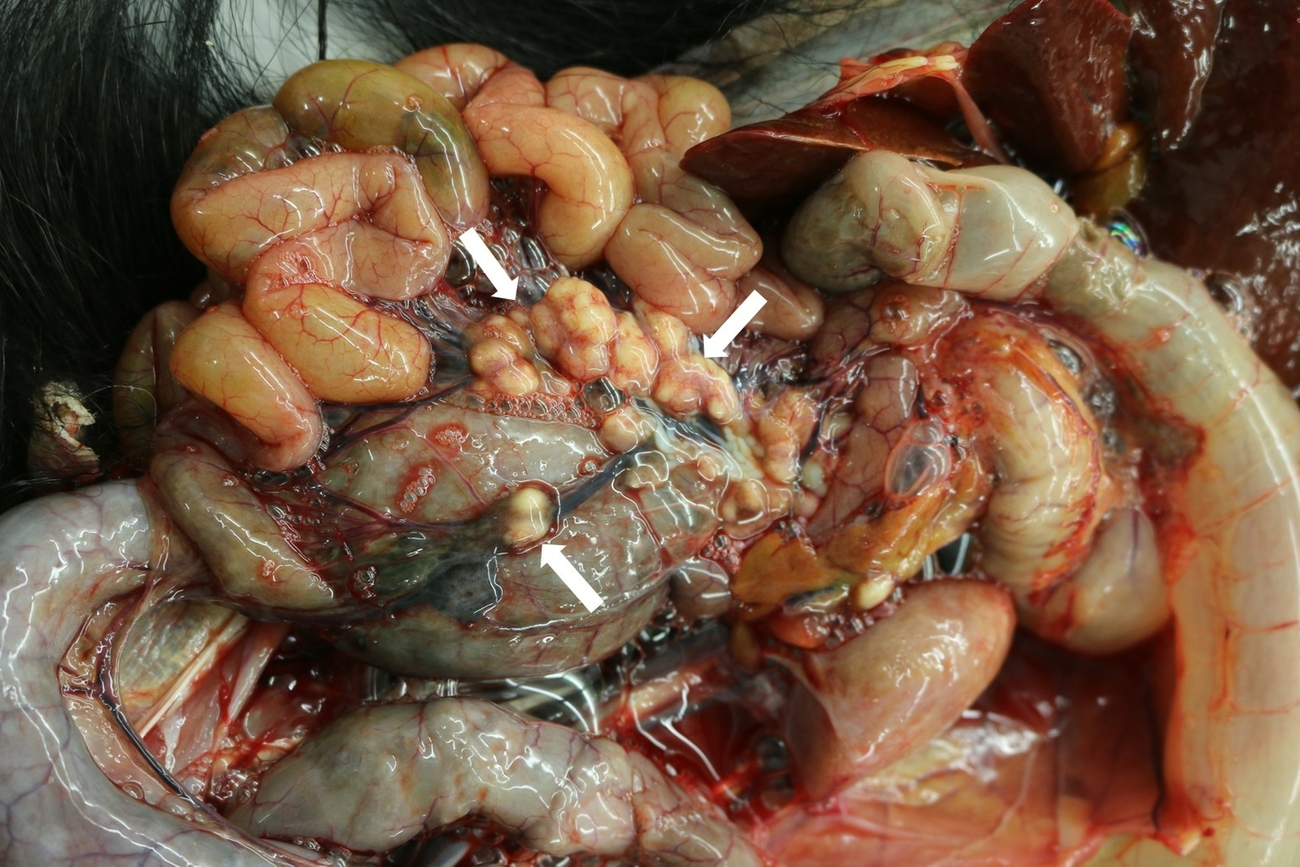
Ultrasonographically, the liver revealed an irregular architecture with hyperechoic regions two months before euthanasia and a hepatic neoplasm was suspected. A blood sample showed a normochromic normocytic anaemia and thrombocytopenia as well as azotemia (urea nitrogen 58 mg/dl [Reference (median) serum values (ISIS/ZIMS): 11 mg/dl], creatinine 1.1 mg/dl [0.5 mg/dl]) and elevated liver (AST 154 U/l [49 U/l], Gamma-GT 724 U/l [29 U/l]) and pancreatic enzymes (α-amylase 683 U/l [103 U/l]) (Species 360 ZIMS 2021). Due to clinical deterioration the animal was euthanized. At necropsy, the cachectic animal revealed multifocal to coalescing, up to 2.0 cm in diameter large granulomas in myocardium, liver, pancreas, spleen, mesenterium, pulmonary, hepatic, intestinal and renal lymph nodes (Fig. 2).
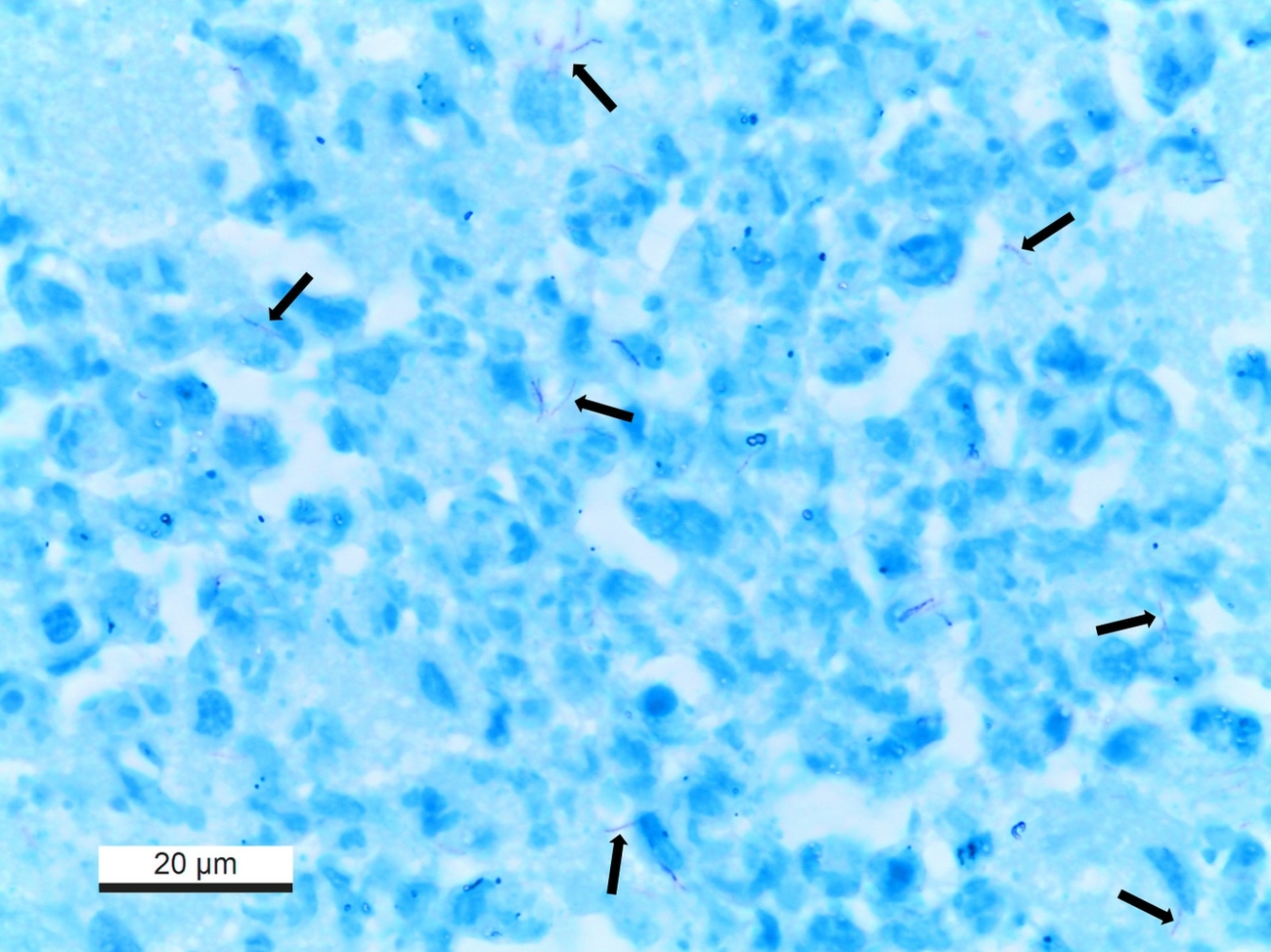
Histologically, lymph nodes and affected parenchymatous organs contained multifocal to coalescing granulomatous to necrotizing inflammation with dystrophic mineralisation. Granulomas showed no fibrous demarcation. Langhans giant cells were not observed. The left cheek had severe ulcerative necrotizing dermatitis with focal histiocytic infiltration in the subcutis. Using Ziehl-Neelsen’s stain few, beaded, elongated acid-fast bacilli were detectable in all lesions mentioned above (Fig. 3). Additionally, the liver revealed moderate bile duct proliferations and multifocal hemosiderin storage detected with Perls’ Prussian blue stain within macrophages. Neoplastic lesions were not detected. In the lung, a mild histiocytic interstitial pneumonia and anthracosis but no granulomas were present. The kidneys were affected by a moderate mesangial proliferative glomerulonephritis and pyelonephritis.
Bacteriological and molecular genetic identification, molecular fine typing
For bacteriological culture, approximately 1 g of liver and pulmonary lymph node were homogenized, decontaminated (1% NALC-NaOH) and subsequently cultivated on Stonebrink/PACT and Löwenstein-Jensen/PACT agar slants as well as in Kirchner bouillon (all media Artelt-Enclit GmbH, Germany). The media were incubated at 37°C and checked weekly for bacterial growth. After four weeks grown mycobacteria were differentiated by DNA sequencing 16S rRNA, the hsp65 as well rpoB genes (Telenti et al 1993, Kirschner and Böttger 1998, Adékambi et al. 2003). The DNA sequences yielded were analysed using NCBI BLASTn with a limited search to type material (Altschul et al. 1997). Isolates from liver and pulmonary lymph node were both identified as M. xenopi.
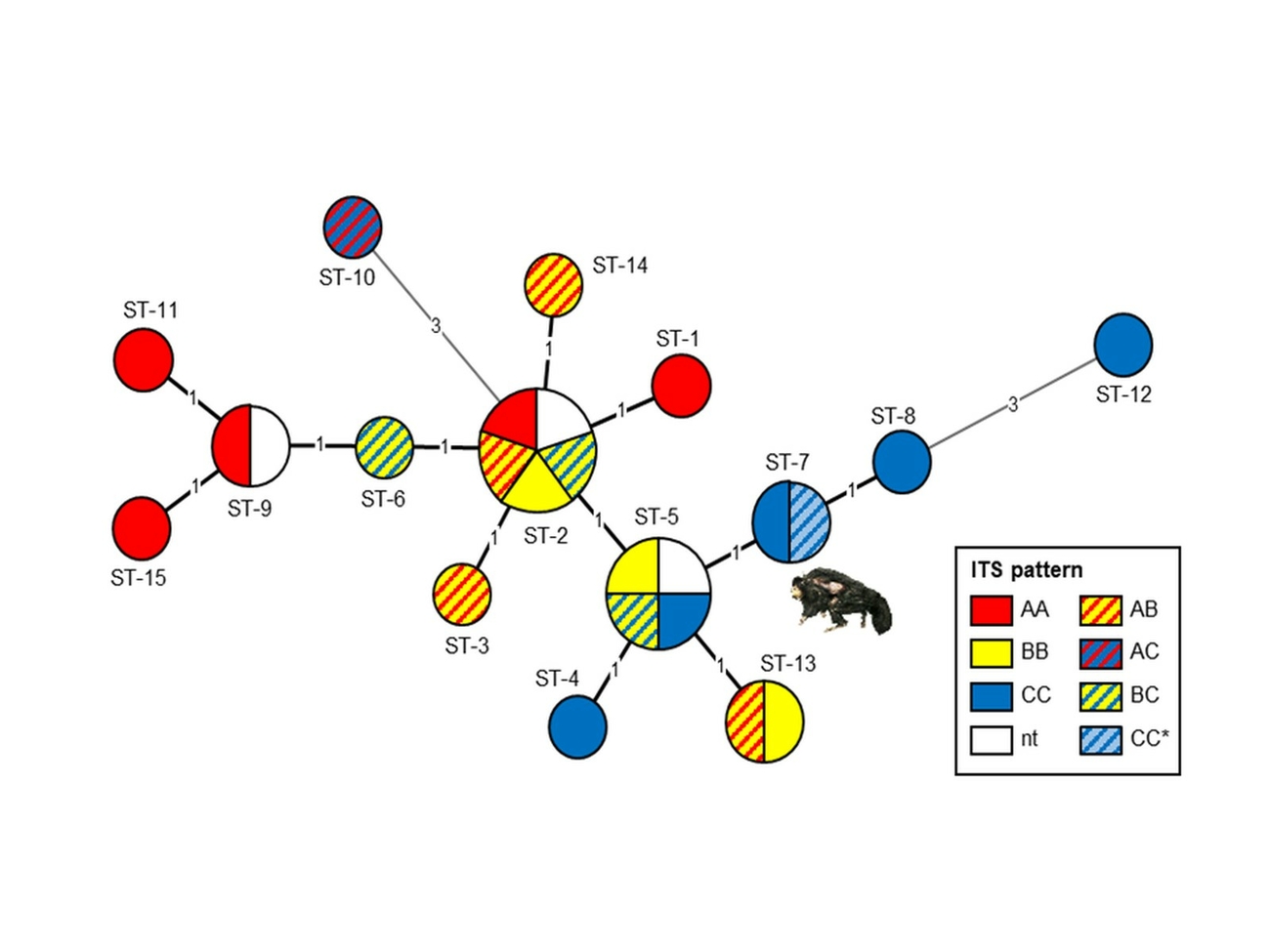
For molecular fine typing of both isolates a multi locus sequence typing (MLST) analysis and internal transcribed spacer (ITS) sequevar identification was performed according to published protocols (Roth et al. 1998, Alexander et al. 2014). In brief, for MLST DNA fragments of the house keeping genes atpD, fusA1, glnA1, pheT, secA1, and topA were amplified, purified, and double stranded nucleotide sequenced (Eurofins Genomics, Germany). To identify the ITS sequevar the 16S-23S ITS region was amplified by PCR, purified and nucleotide sequenced. The SNP analysis and comparison of the sequences were done using Geneious Prime® 2019.2.3 (Biomatters Ltd., New Zealand), the construction of the minimum spanning tree using BioNumerics® 7.6.2 (Applied Maths, Belgium). In both isolates allele Mxe-C was present, but during the doubled stranded sequencing it became obvious that the isolates possess two rRNA operons, differing in one thymine deletion at position 1,449 of the 16S rRNA gene. Nevertheless, in both rRNA operons allele Mxe-C were present. As both isolates possessed the identical thymine deletion only one isolate (from the lymph node) was analysed by MLST. The German saki isolate was assigned to MLST type ST7, a type already described by Alexander and colleagues in human M. xenopi isolates from Canada (Alexander et al. 2014).
Discussion
M. xenopi is a non-tuberculous, slowly growing, thermophilic, Runyon group II mycobacterial species that was first isolated from skin lesions of Xenopus laevis, an African frog (Schwabacher 1959). Although M. xenopi has been isolated from a broad spectrum of animal species (pig, cattle, bird, cat, ferret), it seems to be of minor relevance in veterinary medicine. This is in accordance with the experience of the German National Reference Centre for Mycobacteria in Borstel (personal information) and the German National Reference Laboratory for Bovine Tuberculosis, Friedrich-Loeffler-Institute, where none, respectively only three M. xenopi isolates from animals were identified over the last years. Experimental infections in guinea pigs, rabbits, frogs and chickens resulted in granulomas at the inoculation site and even after several months, no dissemination was observed (Schwabacher 1959). This is contrast to reports of natural infections in cats (MacWilliams et al. 1998, Meeks et al. 2008), a ferret (Davendralingam et al. 2017) and a conure bird (St-Jean et al. 2018) with disseminated granulomatous lesions.
In humans M. xenopi was the third most frequently isolated species in a worldwide surveillance of human NTM lung disease in 2008 (Hoefsloot et al. 2013). Infections are mainly opportunistic in immunocompromised or concurrently infected patients resulting in lung disease, lymphadenitis, skin and soft tissue infection and disseminated forms (Becovier and Vincent 2001).
In the case described here, a disseminated M. xenopi infection was found, however, the portal of entry could not be determined. The infection and clinical course may have been triggered by the deslorelin implant. Due to the hormonal suppression the saki was probably exposed to increased subtle stress within the animal group especially with two adult sons without deslorelin chip in the same enclosure. M. xenopi infections are often related to water-supply systems (Becovier and Vincent 2001) or peat (Trckova et al. 2006). Both sources of infection are conceivable in the enclosure of the sakis; especially as different kinds of bedding and enrichment materials were used in the indoor enclosures. Furthermore, it has to be considered that the relatively high temperature of the indoor enclosure (up to 22°C) might have favoured the growth of the thermophilic bacterium (Torkko et al. 2000). Other animals in the same enclosure could potentially be suspected as possible sources of infection, but mycobacterial infection was not found in any of these animals.
Beside airborne and alimentary infection, percutaneous infection resulting in the remaining dermal ulcerative mycobacteriosis would be another possible route of infection. Absence of granulomatous foci in the lungs does not rule out air-born infection in the sense of an incomplete primary complex. Pulmonary and intestinal lymph nodes and parenchymatous organs showed chronic granulomatous inflammations without fibrous demarcation which is typical for tuberculosis in non-human primates and similar to observations in the M. intracellulare infected white-faced saki (Heard et al. 1997). This finding together with the lack of epitheloid macrophages and giant cells was also reported in atypical mycobacteriosis of black-bearded sakis (Murai et al. 2012) and was interpreted as an inappropriate immune response favouring disseminated mycobacterial infection. The low numbers of acid-fast bacilli are characteristic for a paucibacillary mycobacteriosis. The thin, beaded, elongated acid-fast bacilli seem to be typical for M. xenopi (MacWilliams et al. 1998). Genetic fine typing of M. xenopi isolates of the white-faced saki revealed a MLST-type and sequevar combination already known from a human isolate from Canada (Alexander et al. 2014). Preliminary reports indicate an association between M. xenopi genotype and clinical phenotype (Hirama et al. 2018). Unfortunately, further clinical and epidemiological information on the patient from Canada was not available. Currently, 15 MLST types are described (Alexander et al. 2014) and together with 8 Mxe alleles overall 25 different M. xenopi profiles can be distinguished (Fig. 4). Detection of the same MLST pattern and ITS-sequevar in a M. xenopi strain isolated from a non-human primate in Germany and a human patient from Canada indicate a high degree of conservation in the bacterial species. Therefore, a human pathogenic potential of those strains cannot be excluded and should be kept in mind.
Conflict of interest
The authors declare that there is no conflict of interest.
Ethical approval
The authors declare to have met common international ethical guidelines during genesis of the above work, the underlying research and its publication.
Funding
The authors received no financial support for research, authorship, and/or publication of this article.
Authors contribution
Conceptualisation: MP, CO, SB.
Methodology: MP, IM, SB.
Investigation: CO, MP, PW, IM, SB.
Resources: MP, CO, IM.
Original draft preparation: MP, PW, SB.
All authors have read and agreed to the published version of the manuscript.
Address for correspondence
Martin Peters
Chemisches und Veterinäruntersuchungsamt Westfalen
Zur Taubeneiche 10–12
59821 Arnsberg
Martin.Peters@cvua-westfalen.de
References
Adékambi T, Colson P, Drancourt M (2003): rpoB-based identification of nonpigmented and late-pigmenting rapidly growing mycobacteria. J Clin Microbiol 41: 5699–5708. DOI 10.1128/JCM.41.12.5699-5708.2003.
Alexander DC, Marras TK, Ma JH, Mirza S, Liu D, Kus JV, Soualhine H, Escuyer V, Warshauer D, Brode SK, Farrell DJ, Jamieson FB (2014): Multilocus sequence typing of Mycobacterium xenopi. J Clin Microbiol 52: 3973–3977. DOI 10.1128/JCM.01601-14.
Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ (1997): Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25: 3389–3402. DOI 10.1093/nar/25.17.3389.
Bercovier H,Vincent V (2001): Mycobacterial infections in domestic and wild animals due to Mycobacterium marinum, M. fortuitum, M. chelonae, M. porcinum, M. farcinogenes, M. smegmatis, M. scrofulaceum, M. xenopi, M. kansasii, M. simiae and M. genavense. Rev Sci Tech 20: 265–290. DOI 10.20506/rst.20.1.1269.
Davendralingam N, Davagnanam I, Stidworthy MF, Baldrey V, Peters LM, Stapleton N (2017): Transmission of Mycobacterium xenopi to a pet albino ferret (Mustela putorius furo) from a domestic aquarium. Vet Rec: vetrec-2016-104250. DOI 10.1136/vr.104250.
Heard DJ, Ginn PE, Neuwirth L (1997): Mycobacterium avium intracellulare infection in a white-faced saki (Pithecia pithecia). J Zoo Wildl Med 28: 18–188.
Hirama T, Marchand-Austin A, Ma J, Alexander DC, Brode SK, Marras TK, Jamieson FB (2018): Mycobacterium xenopi Genotype Associated with Clinical Phenotype in Lung Disease. Lung 196: 2013–217. DOI 10.1007/s00408-018-0087-9.
Hoefsloot W, van Ingen J, Andrejak C, Ängeby K, Bauriaud R, Bemer P, Beylis N, Boeree MJ, Cacho J, Chihota V, Chimara E, Churchyard G, Cias R, Daza R, Daley CL, Dekhuijzen PN, Domingo D, Drobniewski F, Esteban J, Fauville-Dufaux M, Folkvardsen DB, Gibbons N, Gómez-Mampaso E, Gonzalez R, Hoffmann H, Hsueh PR, Indra A, Jagielski T, Jamieson F, Jankovic M, Jong E, Keane J, Koh WJ, Lange B, Leao S, Macedo R, Mannsåker T, Marras TK, Maugein J, Milburn HJ, Mlinkó T, Morcillo N, Morimoto K, Papaventsis D, Palenque E, Paez-Pena M, Piersimoni C, Polanová M, Rastogi N, Richter E, Ruiz-Serrano MJ, Silva A, da Silva MP, Simsek H, van Soolingen D, Szabó N, Thomson R, Tórtola Fernandez T, Tortoli E, Totten SE, Tyrrell G, Vasankari T, Villar M, Walkiewicz R, Winthrop KL, Wagner D; Nontuberculous Mycobacteria Network European Trials (2013): The geographic diversity of nontuberculous mycobacteria isolated from pulmonary samples: an NTM-NET collaborative study. Eur Respir J 42: 1604–1613. DOI 10.1183/09031936.00149212.
Kirschner P, Böttger EC (1998): Species identification of mycobacteria using rDNA sequencing. Methods Mol Biol 101: 349–361. DOI 10.1385/0-89603-471-2:349.
MacWilliams PS, Whitley N, Moore F (1998): Lymphadenitis and peritonitis caused by Mycobacterium xenopi in a cat. Vet Clin Pathol 27: 50–53. DOI 10.1111/j.1939-165x.1998.tb01015.x.
Mätz-Rensing K, Kaup FJ (2012): Tuberculosis in Nonhuman Primates – the disease. https://www.dpz.eu/en/unit/pathology/research/pathology-of-non-human-pr… (letzter Zugriff: 04.11.2021).
Meeks C, Levy JK, Crawford PC, Farina LL, Origgi F, Alleman R, Seddon OM, Salcedo A, Hirsch BJ, Hirsch SG (2008): Chronic Disseminated Mycobacterium xenopi Infection in a Cat with Idiopathic CD41 T Lymphocytopenia. J Vet Intern Med 22: 1043–1047. DOI 10.1111/j.1939-1676.2008.0108.x.
Murai A, Murakami T, Inoue M, Ueda H, Shiihara S, Kimura J, Hirata A, Sakai H, Yanai T (2012): Pathological features of Mycobacterium kansasii infection in black bearded sakis (Chiropotes satanas). J Comp Pathol 147: 566–569. DOI 10.1016/j.jcpa.2012.02.006.
Roth A, Fischer M, Hamid ME, Michalke S, Ludwig W, Mauch H (1998): Differentiation of phylogenetically related slowly growing mycobacteria based on 16S-23S rRNA gene internal transcribed spacer sequences. J Clin Microbiol 36: 139–147. DOI 10.1128/JCM.36.1.139-147.1998.
Schwabacher H (1959): A strain of Mycobacterium isolated from skin lesions of a cold-blooded animal, Xenopus laevis, and its relation to atypical acid-fast bacilli occurring in man. J Hyg (Lond) 57: 57–67. DOI 10.1017/s0022172400019896.
St-Jean G, Gagnon CA, Soualhine H, Tremblay M, Beaulieu AA, Sylvestre D (2018): Mycobacterium xenopi systemic infection in a domestic fieryshouldered conure bird (Pyrrhura egregia). JMM Case Rep 5: e005158. DOI 10.1099/jmmcr.0.005158.
Telenti A, Marchesi F, Balz M, Bally F, Böttger EC, Bodmer T (1993): Rapid identification of mycobacteria to the species level by polymerase chain reaction and restriction enzyme analysis. J Clin Microbiol 31: 175–178. DOI 10.1128/jcm.31.2.175-178.1993.
Torkko P, Suomalainen S, Iivanainen E, Suutari M, Tortoli E, Paulin L, Katila ML (2000): Mycobacterium xenopi and related organisms isolated from stream waters in Finland and description of Mycobacterium botniense sp. nov. Int J Syst Evol Microbiol 50: 283–289.
Trckova M, Zraly Z, Bejcek P, Matlova L, Beran V, Horvathova A, Faldyna M, Moravkova M, Shitaye JE, Svobodova J, Pavlik I (2006): Effect of feeding treated peat as a supplement to newborn piglets on the growth, health status and occurrence of conditionally pathogenic mycobacteria. Vet Med (Praha) 12: 544–554. DOI 10.17221/5589-Vetmed.
Species 360 ZIMS (2021): Version 2.25.5. Tests and Results Report for Taxonomic Grouping Pithecia pithecia / White Faced Saki, median values (colorimetry, serum) for urea nitrogen, creatinin, AST, y-GT, amylase, report date: 8th August 2021.


Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2021-24-Peters.pdf (0.22 MB) herunterladen möchten


{kind=link}
{kind=link}
{kind=link}