
Coronaviren in Fledertieren
Berliner und Münchener Tierärztliche Wochenschrift 134, 1-16
DOI: 10.2376/1439-0299-2020-44
© Schlütersche Fachmedien GmbH. 2021
Publiziert: 05/2021
Summary
This article summarizes current detections of coronaviruses in bats in different geographic regions worldwide, and underlines the potential role of bats as ancestor and reservoir hosts of different coronaviruses. Bats can be found in almost all global habitats with the notable exception of the high arctic and a few isolated oceanic islands. First, we give an overview on the potential role of bats as carriers of zoonotic viruses, underlining the relevance of specific physiological traits of representatives of this mammalian order, that are supporting the transmission of these disease agents. Research in this field was predominantly initiated by the SARS coronavirus event in 2003, and in the meantime, bats are assumed to be the ancestor hosts for numerous alpha and beta coronaviruses. Bat associated coronaviruses have been detected on all continents where bats are distributed. Since the beginning of the COVID-19 pandemic, coronavirus research has again been considerably intensified, leading to a gain of knowledge regarding the interaction between bats and coronaviruses that will be valuable in the management of potential future outbreaks originating from bats. Finally, an outlook is given on research gaps that could be invaluable when dealing with future pandemic events.
Zusammenfassung
Dieser Artikel gibt einen Überblick über die bisherigen Nachweise von Coronaviren in Fledertieren in verschiedenen geografischen Regionen weltweit. Darüber hinaus wird auch die potenzielle Rolle der Fledertiere als Ursprungs- und Reservoirwirt für die verschiedenen Coronaviren erörtert. Bemerkenswerterweise kommen Fledertiere weltweit in fast allen Habitaten vor, mit Ausnahme der hohen Arktis und einiger isolierter Ozeaninseln. Zunächst wird ein Überblick über die Bedeutung der Fledertiere als potenzielle Überträger von Viren mit zoonotischem Potenzial gegeben und dabei die Bedeutung der speziellen physiologischen Charakteristika dieser Säugetier-Ordnung hervorgehoben, die die Verbreitung solcher Erreger unterstützen. Die Forschung auf diesem Gebiet wurde erst durch das SARS-Coronavirus-Geschehen 2003 angestoßen, wobei derzeit davon ausgegangen wird, dass Fledertiere als Ursprungswirt für zahlreiche alpha- und beta-Coronaviren eine bedeutende Rolle spielen. Auf allen Kontinenten mit Fledertiervorkommen wurden inzwischen auch Fledertier-assoziierte Coronaviren nachgewiesen. Seit dem Beginn der COVID-19 Pandemie wurden die Arbeiten nochmals deutlich intensiviert, wodurch ein Wissensgewinn in Bezug auf die Interaktion zwischen Fledertieren und Coronaviren erreicht wurde, der auch zukünftig bei der Bekämpfung pandemischer, möglicherweise von Fledertieren ausgehender Infektionsgeschehen von Nutzen sein wird. Schließlich wird ein Ausblick auf noch offene Forschungsfragen gegeben, deren Beantwortung beim Umgang mit zukünftigen pandemischen Ereignissen von Nutzen sein könnten.
Introduction
This article summarizes the currently known detections of coronaviruses in bats in different geographic regions worldwide, and underlines the potential role of bats as ancestor and reservoir hosts for different coronaviruses. This information therefore complements what has been summarized in other recently published review articles regarding different aspects of infectiology, bat immunology, virus/bat interactions and bat ecology (Banerjee et al. 2019, 2020, Gorbunova et al. 2020, Irving et al. 2021, Letko et al. 2020). It includes a short insight into published in vitro and in vivo work, followed by a discussion of public health relevance of these findings, as well as the research gaps in this field.
Bats as potential reservoirs for zoonotic diseases agents
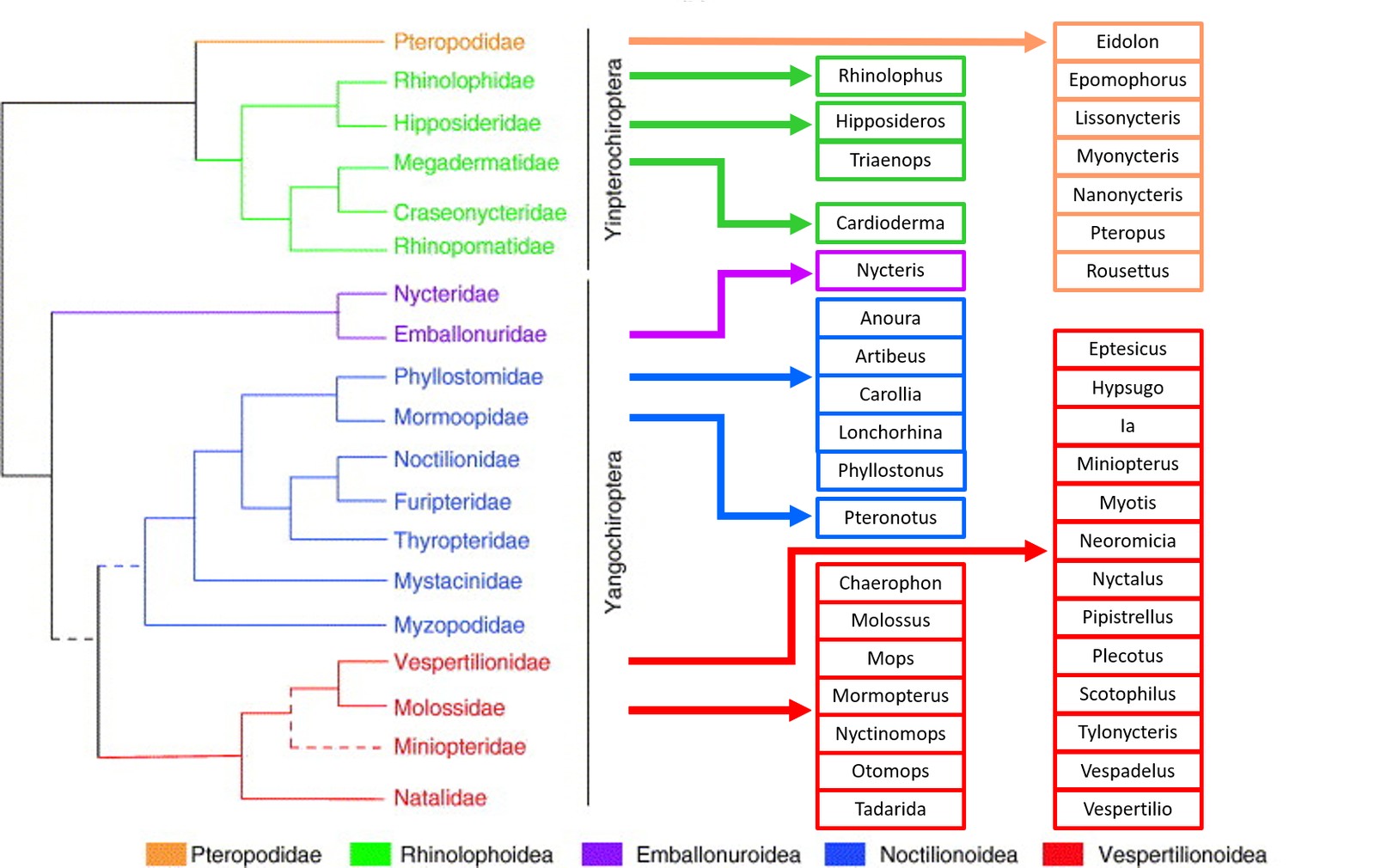
In recent years, bats have increasingly moved into public and scientific attention after they have been identified as reservoir hosts of a number of disease agents. Bats form the second largest order of mammals aside from rodents, with more than 1300-1400 known species. This already partly explains their role as reservoir hosts for a number of relevant viruses, a perception which has been supported by a recent article discussing the relevance of mammals and birds as possible reservoir hosts (Mollentze and Streicker 2020). Following morphological, behavioural and molecular characteristics, the order chiroptera divides into Yinpterochiroptera (comprising the family Pteropodidae (fruit bats, formerly known as megabats), and the Rhinolophoidae superfamily with 5 families) and Yangochiroptera (comprising the remaining 14 bat families, that all use laryngeal echolocation). In general, bats exist in highly diverse habitats on six continents, but with the notable absence in the high Arctic, the Antarctic and a few isolated oceanic islands (Fenton 2015). The different bat families and species display considerable differences in their habitat, lifestyle, diet and social structure, resulting in large variations in the potential risk they may pose to public and veterinary health. In general, bats harbour specific life traits that increases their relevance as virus reservoirs: Bats are the only flying mammals and are thus able to cover large distances within short periods of time, with the annual migration of some species covering up to 1500 kilometres (Thomas 1983). Bats are also very gregarious and tend to form large colonies of up to tens of thousands of individuals, increasing the risk of virus spread through the colony. Some bat species even share their roosts with other animal species or roost close to humans, (e.g. in roofs of buildings) increasing the risk for a spillover infection (Kemenesi et al. 2015, Steyer et al. 2013). Interestingly, their body temperature can span a wide range between 10°C during torpor and hibernation, up to 43°C in hot roosts (Bronrier et al. 1999, Maloney et al. 1999) or during flight. It has been postulated that the elevated body temperature during flight activity (‚flight as fever‘) may mimic the fever reaction of other mammals, thereby activating the immune responses (O’Shea et al. 2014). On the other hand, the activity of the immune system is downregulated during the torpor phases (torpor bouts during summer and torpor during hibernation) of temperate zone bats, which may hamper virus clearance. Finally, by mechanisms that are still not fully understood, their immune system is able to protect them from developing disease after infection with a large number of viruses that are highly pathogenic to most other mammalian species. The current knowledge of the interaction of viral pathogens with the bat immune system has been summarized in a number of recent reviews (Banerjee et al. 2020, Gorbunova et al. 2020, Letko et al. 2020).
Top Job:
Many zoonotic viruses from bats may carry high pathogenic potential for humans, livestock or companion animals, especially in bats of tropical regions. The most prominent examples are filoviruses that are highly pathogenic to humans (e.g. Marburg and Ebola virus), henipaviruses that are highly pathogenic to pigs, horses and humans (e.g. Nipah and Hendra virus), lyssaviruses that are fatal to livestock, companion animals and humans (e.g. Rabies and Lagos Bat virus) as well as coronaviruses that are pathogenic to livestock, companion animals and humans (SARS-CoV, MERS-CoV and SARS-CoV-2) (Calisher et al. 2006, Moratelli and Calisher 2015, Nabi et al. 2021). In general, bats are capable of propagating and shedding viruses of all Baltimore classifications (Chen et al. 2014, Hayman 2016). Moreover, bats have been postulated as ancestral hosts for the most relevant mammalian paramyxoviruses (Drexler et al. 2012), alpha- and beta-coronaviruses (Woo et al. 2012) as well as lyssaviruses (Badrane and Tordo 2001, Delmas et al. 2008).
However, despite the abundance of viruses detected in bats, spillover events are still relatively rare, as viruses need to overcome a number of barriers imposed by ecological, geographical and social factors. This leads to variable levels of pathogen pressure, probability of virus survival in the environment, probability of exposure, and finally probability of infection of the host. These spillover cascades have been comprehensively summarized in two articles published by Plowright and colleagues (Plowright et al. 2015, 2017).
Coronaviruses (CoVs)
As classified by the International Committee on Taxonomy of Viruses (ICTV, 2019) Coronaviridae form the largest family in the order Nidovirales with the highest relevance to human and animal health. They harbour a single-stranded positive-sense RNA genome, ranging among the largest RNA virus genomes. The family is split into two subfamilies, the first of which, Letovirinae contains only one known species, Microhyla letovirus 1, which infects amphibia. The other subfamily, Orthocoronavirinae, comprises four genera: alpha-, beta-, gamma- and delta-CoVs. While alpha- and beta-CoVs infect mammals, infections with gamma- and delta-CoVs are mainly confined to birds. Therefore, in the following, we focus our review on infections of bats with alpha- or beta-CoVs. Interestingly, in each subgenus of both alpha- and beta-CoV genera, at least one virus species has been found in bats, highlighting the relevance of bats as hosts for CoVs, and again pointing to a coevolution of bats with these two genera (Woo et al. 2012).
Human CoV infections are of high importance to public health, with the emergence of SARS-CoV in 2003 (Drosten et al. 2003), and MERS-CoV in 2012
(Zaki et al. 2012) causing significant outbreaks. The most notable human coronavirus is SARS-CoV-2 which emerged in 2019 (World Health Organization 2020b) leading to an unprecedented pandemic. Aside from the new emergence of CoV species, others with relatively low pathogenicity play a key role in seasonal respiratory infections with global distribution. The most relevant virus species are hCoV NL63, hCoV 229E (alpha CoV) and hCoV OC43, hCoV HKU1 (beta CoV). On the other hand, a number of CoV infections, addressed in detail in other articles of this Special Issue, have a strong relevance for veterinary medicine.
Coronavirus infections in bats
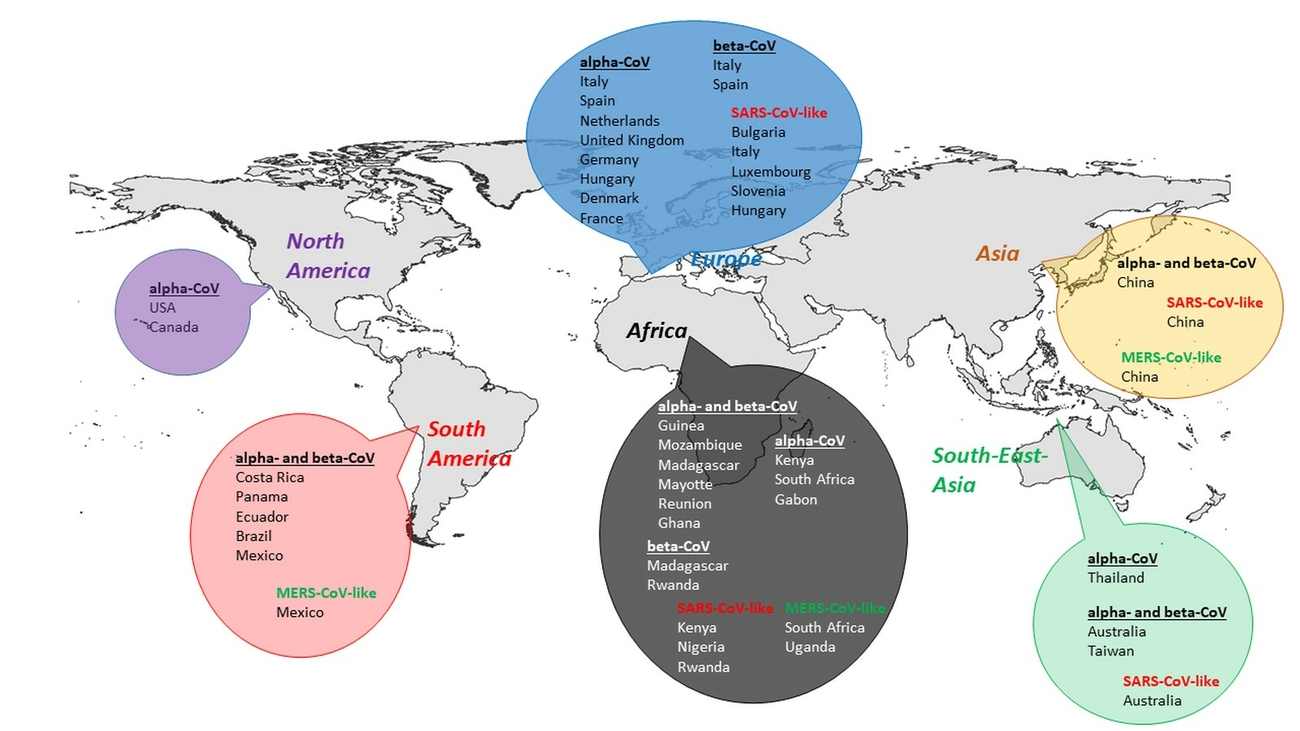
Prior to the SARS-CoV epidemic, bats were not known to be hosts for CoVs. Since then they have been found to carry many more CoVs, with 30 complete genomes sequenced, and many more partial genome sequences (Wong et al. 2019). They were shown to be the group of mammals harbouring the largest number of CoVs (Fig. 1). To date, only alpha- and beta-CoVs have been detected in bats which have been identified in Asia, Europe, Africa, North and South America, as well as Australasia. With higher detection rates, alpha-CoVs appear more widely distributed than beta-CoVs (Wong et al. 2019). Bats are hosts for many alpha- and beta-CoVs (Shi 2013, Woo et al. 2012) and phylogenetic analysis suggest, that bat-CoVs may be the ancestors for many CoVs in other mammals (Wong et al. 2019).
In general, bat CoVs are characterized by a distinct species-, genus- or family-specificity, while showing a wide geographic distribution across continents
(Chu et al. 2006, Drexler et al. 2014, Gloza-Rausch et al. 2008, Pfefferle et al. 2009, Poon et al. 2005, Tang et al. 2006, Woo et al. 2006a). Interestingly, the diversity of bat related CoVs seems to be correlated with the diversity of bat species in a certain geographical region (Anthony et al. 2017b). It could be shown that the
same alpha-CoV (Miniopterus bat coronavirus HKU8), which had previously been reported in Miniopterus species in China and Hong Kong, occurred in M. schreibersii in Bulgaria, over 8000 km away (Drexler et al. 2010).
In the following, we summarize the current knowledge regarding alpha- and beta-CoVs in bats in the different geographical regions and climatic zones worldwide (Tab. 1). Detection of CoV-related RNA has been most successful from faecal samples in all previous studies, indicating a tropism to the digestive system in
Chiroptera (Anthony et al. 2013, Pfefferle et al. 2009, Tang et al. 2006). We include an extensive list of references for further reading where the detailed information goes beyond the scope of this short review article.
Bat-related CoV livestock and companion animals
As mentioned before, bats have been postulated as ancestor hosts for a number of relevant alpha- and beta-CoVs. Moreover, a common ancestor has been postulated for most relevant CoVs in veterinary medicine, namely TGEV (transmissible gastroenteritis virus) in pigs, CCoV (canine coronavirus) in dogs and FCoV (feline coronavirus) in cats (Le Poder 2011). Although the host species of this ancestor virus has not been identified yet, it may have been bat-related. In 2017, a novel HKU2-related CoV, later reported as SADS-CoV (Swine acute diarrhoea syndrome coronavirus, or porcine enteric alphacoronavirus), caused a fatal diarrhoea syndrome in pigs in China and resulted in the loss of almost 25.000 piglets (Gong et al. 2017, Zhou et al. 2018). SADS-CoV shows a 95% sequence homology to a bat-related HKU2-CoV, that has been detected in Rhinolophus bats in Southern China in 2007 (Lau et al. 2007).
Coronaviruses in temperate bats
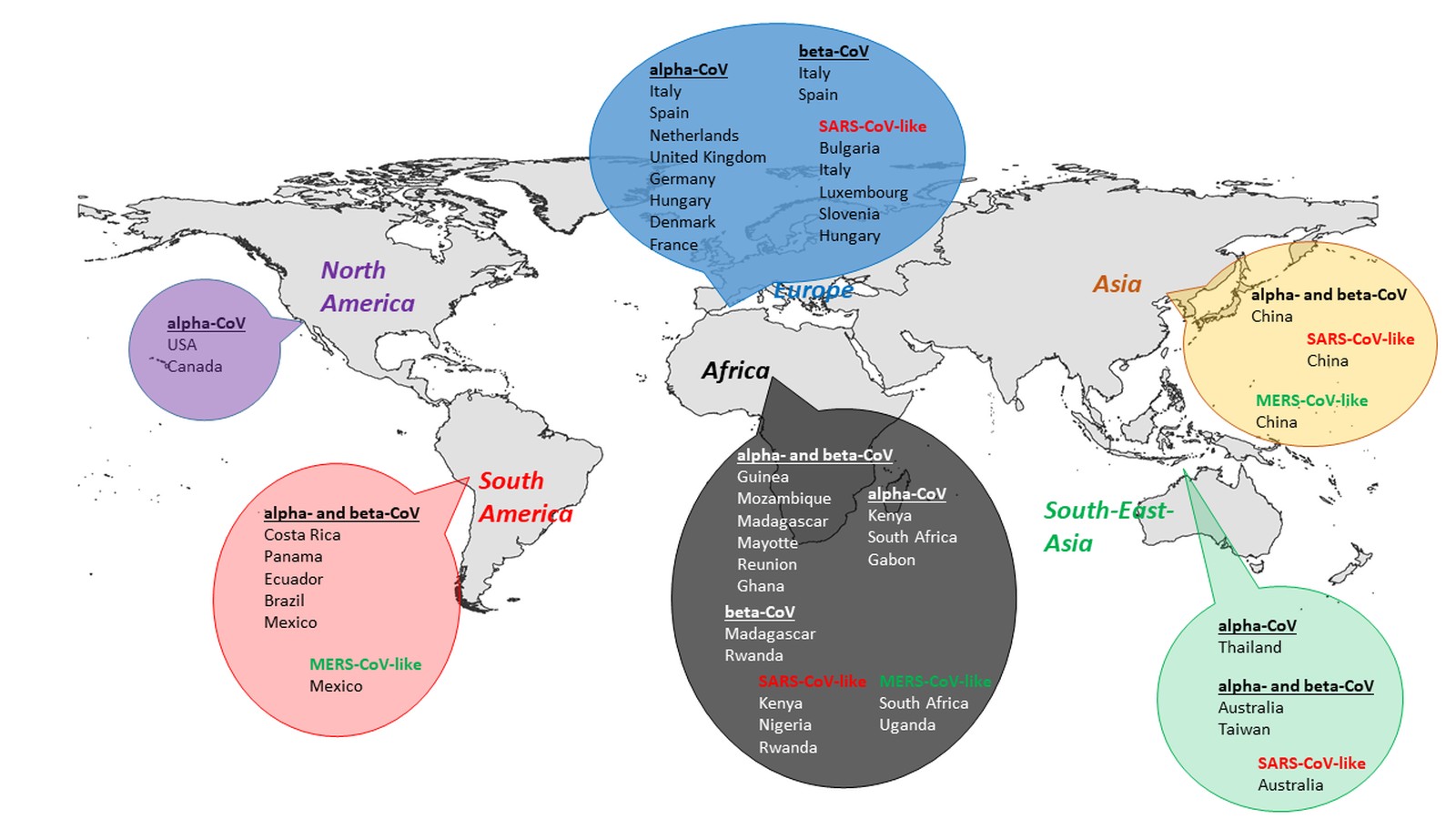
Europe
Three alpha-CoVs, fully sequenced by next generation sequencing, were detected in Pipistrellus kuhlii bats in Italy, of which one strain showed a high level of sequence similarity to a Chinese bat-CoV (De Sabato et al. 2019). In another study in Italy, alpha- and beta-CoVs were detected using a pan-CoV-PCR assay in different vespertilionid and rhinolophoid bat species, with Pipistrellus kuhlii bats carrying both, alpha- and beta-CoVs (Lelli et al. 2013). In Spain, 14 alpha- and beta-CoVs were detected in nine bat species (Falcón et al. 2011) and in the Netherlands the overall estimated prevalence of CoVs in Myotis, Nyctalus and Pipistrellus bats was 16.9% (Reusken et al. 2010). In 2012 the first CoVs were detected in bats in the United Kingdom by PCR, with particularly high prevalence in Myotis nattereri, which have been found living in close proximity to humans (August et al. 2012). Testing of 315 bats in northern Germany by PCR revealed four lineages of alpha-CoVs in Myotis and Pipistrellus bats with a overall prevalence of 9.8% (Gloza-Rausch et al. 2008). In another study the average detection rate for CoVs in Myotis and Pipistrellus bats by PCR was only 1.4% of 957 analysed samples from different regions in Germany (Fischer et al. 2016). In Hungary a detection rate of 1.79% was found in Myotis, Pipistrellus and Rhinolophus bats, testing 447 samples from 24 bat species (Kemenesi et al. 2014). Alpha-CoVs were also detected by PCR in Myotis, Pipistrellus and Eptesicus bats in Denmark, of which some are showing a close resemblance to coronavirus sequences obtained from bats in Germany and the UK (Lazov et al. 2018). Twelve out of 504 bat intestinal samples from France were tested positive by PCR, revealing alpha-CoVs in Myotis, Pipistrellus and Miniopterus bats with high sequence similarities to CoV sequences obtained from other European countries such as Germany, Luxembourg, Hungary and Bulgaria (Monchatre-Leroy et al. 2017).
A study on Rhinolophids in Bulgaria showed, that these bats carried SARS-related beta-CoVs at high frequencies and RNA levels (26% of 389 animals were positive; up to 2.4 × 108 copies per gram of feces) (Drexler et al. 2010). In Italy (Balboni et al. 2011, 2012, Lecis et al. 2019), Luxembourg (Pauly et al. 2017), Slovenia (Rihtarič et al. 2010) and Hungary (Kemenesi et al. 2014) SARS-like CoVs were detected by PCR in Rhinolophus bats. In another study, beta-CoV sequences obtained from Pipistrellus bats in Romania and Ukraine showed relatedness to hCoV EMC/2012 (Annan et al. 2013), while MERS-like CoVs were detected in Eptesicus, Pipistrellus, Nyctalus and Hypsugo bats from Italy (De Benedictis et al. 2014, Lelli et al. 2013, Moreno et al. 2017), with two complete CoV genome sequences obtained from a Pipistrellus khulii and Hypsugo savii bat (Moreno et al. 2017).
North America
To allow a science-based risk assessment regarding CoV infections of bats in North America, a small number of bats of seven Rocky Mountain vespertilionid bat species were screened for CoVs by PCR. In 17% of fecal samples of big brown bats (Eptesicus fuscus) and 50% of Myotis occultus alpha-CoVs were detected (Dominguez et al. 2007). In a larger follow-up study, more than 1000 samples collected from 17 bat species in Colorado were analysed, with alpha-CoVs detected in up to 10% of fecal samples of four species. The highest detection rate was seen in E. fuscus. The authors concluded, that given the exclusive detection of alpha-CoVs in these bats, there appears to be a much more limited CoV diversity in New World bat species than in Old World bat species (Osborne et al. 2011). A later study was consistent with the restriction of North-American bat populations to infections with alpha-CoVs by PCR analysis of fecal samples of two out of 19 Brazilian free-tailed bats (Tadarida brasiliensis) in Florida (Bonny et al. 2017). A study summarizing the analysis of 31 vespertilionid bats also confirmed the presence of alpha-CoVs in these species in Canada (Misra et al. 2009). Persistent infections with Chiroptera-alpha-CoVs were detected in the intestines and lungs of some North American little brown bats (Myotis lucifugus) after four months of hibernation (Subudhi et al. 2017, 2019).
Asia
After the SARS-CoV epidemic, CoV research in bats intensified, and a large number of studies was performed in Asia. In 2005, two independent teams reported the discovery of SARS-like CoVs (SL-CoVs) in bats and suggested that bats are natural reservoirs of SARS-CoV (Lau et al. 2005, Li et al. 2005). Of the various bat species, horseshoe bats (Rhinolophus spp.) were suggested as a reservoir of SARS-CoV.
While the previously known human CoVs (HCoV-229E, HCoV-OC43, HCoV-HKU1 and HCoV-NL63) usually cause mild symptoms, the more recently discovered human CoVs have higher virulence and as a result have a more significant public health impact. The emergence of the beta-CoV SARS-CoV occurred in 2003 after a possible transmission from civet cats or raccoon dogs as an intermediate host to humans (Peiris et al. 2003). Within a few months, SARS-CoV infected around 8000 people in more than 30 countries with a lethality rate of ~10% (Peiris et al. 2003). Another beta-CoV, MERS-CoV emerged in 2012, causing more than 2500 cases in 27 countries with a lethality rate of 35% (World Health Organization 2019, Zaki et al. 2012, Zumla et al. 2015). Dromedary camels seem to represent a natural reservoir of MERS-CoV (Killerby et al. 2020, Lau et al. 2017, Paden et al. 2018, Wernery et al. 2017), but again bats have been postulated as the original reservoir host (Killerby et al. 2020), as several closely related CoVs were detected in different bat species (Corman et al. 2014, Hu et al. 2015). A recent PCR-based study testing more than 1000 bats from >30 different species in South China revealed 89 different beta-CoV strains from eight vespertilionid bat species, of which a number of sequences showed high similarities to MERS-CoV (Luo et al. 2018). Two full-length sequences indicate a natural recombination event between this bat MERS-related CoV and another bat coronavirus, HKU4. Moreover, this study expands the known host range of MERS-related CoVs and confirms the geographical distribution of MERS-like CoVs outside the Old World and North America.
A screening survey of bat intestinal samples collected from 21 different bat species in the Tibet Autonomous Region revealed an overall CoV positivity rate of more than 5%. Interestingly, 84% of these sequences belonged to beta-CoVs that are generally less frequently reported in bats, while the remaining 16% were related to alpha-CoVs (Xu et al. 2016).
Next, the HKU2-related SADS-CoV has frequently been described in Rhinolophus bats in China before the first SADS outbreaks in Chinese pig farms were reported (Gong et al. 2017, Zhou et al. 2018). The current CoV findings in Chinese bats have been comprehensively summarized by (Fan et al. 2019).
Then in December 2019, a novel SARS-CoV, later designated SARS-CoV-2, was first detected in Wuhan, China, and was possibly epidemiologically associated to a seafood market (Zhu et al. 2020). SARS-CoV-2 represents the seventh CoV to be identified in humans since the 1960s (Su et al. 2016, Ye et al. 2020). The recent SARS-CoV-2 outbreak has an estimated case-fatality rate lower than that seen during the SARS-CoV outbreak, but its distribution and speed of its spread resulted in a much higher disease burden (He et al. 2020). By March 11th 2020, the WHO declared the ongoing outbreak a pandemic (World Health Organization 2020a). To date (April 2, 2021), 130 million COVID19-cases have been reported worldwide, with more than 2.8 million fatalities (Johns Hopkins University – Coronavirus Resource Center 2020). All seven HCoVs are zoonotic viruses with likely evolutionary origins in bats (Corman et al. 2018, Huynh et al. 2012, Pfefferle et al. 2009, Vijaykrishna et al. 2007). Whilst some have been directly transmitted from bats to humans, for other CoVs an intermediate host was implicated, for example, camels for MERS-CoV or civets for SARS-CoV (Corman et al. 2014, Guan et al. 2003, Lau et al. 2005, Li et al. 2005, Luo et al. 2018). For SARS-CoV-2, the closest relative (RaTG13) has been identified in a Rhinolophus affinis bat from China with 96.2% sequence homology to SARS-CoV-2. Therefore, bats have been also postulated as the natural reservoir for this virus (Guo et al. 2020, Zhou et al. 2020). The role of an intermediate species, is still not fully understood (Lam et al. 2020, Zhou et al. 2020).
Coronaviruses in tropical and subtropical bats
Africa
It was shown in a number of studies that insectivorous and frugivorous bats across Africa harbour a diverse number of CoVs. Antibodies against SARS-CoV antigen were detected in 6.7% of 705 bat serum specimens comprising 26 species collected in South Africa and the Democratic Republic of Congo (DRC) (Müller et al. 2007). In 11% of 319 tested bats from 14 genera in Guinea, numerous different alpha- and beta-CoVs were found using PCR (Lacroix et al. 2020). Testing of bats from Mozambique, Madagascar, Mayotte and Reunion Island revealed a large diversity of alpha- and beta-CoVs (Joffrin et al. 2020). Another study showed a significant diversity of CoVs circulating in bats in Kenya, which are phylogenetically distant from any previously described human and animal CoV (Tao et al. 2012). In Pteropus and Eidolon bats in Madagascar 14 CoVs were detected, forming distinct clusters within the beta-CoVs (Razanajatovo et al. 2015) and in Rwanda new beta-CoVs were recently discovered in Rhinolophus and Hipposideros bats (Markotter et al. 2019, Nziza et al. 2019). Three unique alpha-CoV species were identified in the tissues of the South African bats Neoromicia capensis, Miniopterus spp. and Mops midas, of which the CoV Mops BtCoV1364/SA/11 is closely related to the CoV Chaerephon-BtKY22 from Kenya (Geldenhuys et al. 2013, Tong et al. 2009). The two bat genera, Mops and Chaerephon, are closely related and share an overlapping geographical distribution, sometimes even sharing same roosts, so interspecies transmissions of these alpha-CoVs could easily occur between these two host genera (Geldenhuys et al. 2013). Mops condylurus, in which a new ebolavirus, Bombali virus (BOMV), was recently discovered in Sierra Leone (Goldstein et al. 2018), Kenya (Forbes et al. 2019) and Guinea (Karan et al. 2019), was also shown to harbour alpha-CoVs (Waruhiu et al. 2017).
Some of these described alpha- and beta-CoVs are closely related to human CoVs such as hCoV-229E, SARS- and MERS-CoVs (Lacroix et al. 2020). In Ghana, fecal samples of insectivorous leaf-nosed bats of the genus Hipposideros tested positive for alpha- and beta-CoVs, with the most closely related member of the GhanaBt-CoVGrp1 clade sharing 91.90% nucleotide identity with hCoV-229E in the analysed fragment (Pfefferle et al. 2009). In other studies alpha-CoV sequences obtained from Hipposideros bats in Gabon (Maganga et al. 2020) and Zimbabwe (Bourgarel et al. 2018) grouped with hCoV-229E (Maganga et al. 2020). The evolutionary origin of 229E-related CoVs seem to be in hipposiderid bats (Corman et al. 2015), while sequences closely related to human CoV NL63 were identified in Triaenops bats (Tao et al. 2017). Sequences closely related to SARS-like CoVs were detected in Chaerophon bats in Kenya (Tong et al. 2009) and Hipposideros bats in Nigeria (Quan et al. 2010). Also, in Rwanda, novel beta-CoVs were shown to cluster with SARS-CoV (Nziza et al. 2019). In Nycteris bats in Ghana novel clade 2c betacoronaviruses were detected that are phylogenetically related to the novel hCoV EMC/2012 (Annan et al. 2013). Close relatedness of CoV PML/2011, detected in Neoromicia cf. zuluensis in South Africa, and MERS-CoV support the hypothesis that, like human CoV-229E and SARS-CoV, ancestors of MERS-CoV may exist in Old World insectivorous bats belonging to the family Vespertilionidae, to which the genera Neoromicia and Pipistrellus belong (Ithete et al. 2013). NeoCoV, obtained from a South African Neoromicia capensis bat, and MERS-CoV even belong to one viral species (Corman et al. 2014). Phylogenetic analysis of a MERS-like CoV (strain PREDICT/PDF-2180), detected in a Pipistrellus bat in Uganda, showed that it is closely related to MERS-CoV across much of its genome, further supporting the hypothesis that bats are the evolutionary source of MERS-CoV (Anthony et al. 2017a). In conclusion, there is a large pool of different alpha- and beta-CoVs detected in African bats with presumably many more to be discovered. This is particularly important as some prominent human CoVs seem to originate from CoVs in these bats.
South-East Asia and Australia
A PCR- and serology-based study analysing more than 2000 bats from over 40 different species in the Australasian region revealed a wide-spread presence of CoVs in this area (Smith et al. 2016). Four different CoV genotypes were detected including three potentially new species, whilst no SARS-like beta-CoVs were identified, despite targeting rhinolophid bats. Interestingly, Miniopterus bat coronavirus HKU8 was detected, which had previously been described in Miniopterus bats in China, Hong Kong and Bulgaria, supporting the virus-host co-evolution hypothesis. A recent serological survey of Pteropus poliocephalus bats in Southern Australia revealed, that 42.5% of the analysed 301 sera were reactive to SARS-CoV or a related antigen, providing evidence that these flying foxes had previously been in contact with a SARS like coronavirus, but no reactivity to a MERS-CoV antigen could be demonstrated (Boardman et al. 2020). In another PCR-based study performed on Hipposideros bats in Thailand, bat CoV sequences could be detected that were closely related to those detected earlier in Africa (beta-CoV) or in Europe (alpha-CoV and beta-CoV). This evidence again supports the coevolution theory of bats and CoVs, further supporting the hypothesis of a beta-CoV spillover from Hipposideridae to Rhinolophidae bats, and subsequently from Rhinolophidae to civets and humans (Gouilh et al. 2011).
South America
A pilot study to evaluate the general presence of CoVs in Brazilian bats was performed on > 500 samples from two insectivorous bat species, Molossus molossus and Tadarida brasiliensis, and revealed alpha-CoV sequences in 29 of 150 pooled fecal samples (Lima et al. 2013). Another study revealed alpha- and beta-CoV RNA in 50 of >1500 bats from nine different frugivorous and insectivorous species in Costa Rica, Panama, Ecuador and Brazil, underlining the broad distribution among different bat species (Corman et al. 2013). Meanwhile, alpha- and beta-CoV related RNA was detected in samples from nine frugivorous and four insectivorous bat species in Mexico, with one beta-CoV sequence isolated from an insectivorous bat (Nyctinomops laticaudatus) showing 96.5% identity to MERS-CoV (Anthony et al. 2013), again emphasizing the global distribution of closely related bat-associated CoVs among different bat species.
In summary, the cumulated data in this article show, that CoVs are detectable in all global areas, where bats can be found (Fig. 2). While SARS-like CoVs have not only been detected in Asia, but also in Europe, Africa, and South-East Asia, MERS-like CoVs have so far been identified in Asia, Africa and South America. Moreover, in the temperate zones of Europe, North America, and Asia as well as in the tropical and subtropical zones of South-East Asia and South America, most studies confirmed the presence of CoV lineages, showing relatedness to other already know CoV clades. Some distinct novel alpha and beta-CoVs have been detected in a number of African countries.
Virus isolation from bats and in vivo experiments in bats
While the vast majority of CoV detection in bats is based on molecular detection by PCR, there are comparably few isolates available that allow the analysis of the biological properties of these strains. While attempts to isolate the progenitor viruses of human CoV epidemics were so far unsuccessful for both SARS-CoV and SARS-CoV-2, the isolation of beta-CoVs that are closely related to SARS-CoV in China facilitated in vitro analysis of cell entry and susceptibility (Ge et al. 2013, Yang et al. 2015). Such studies are crucial to gain a better understanding of the biological properties of these agents. However, isolation of CoVs from free-ranging bats does not allow a profound analysis of the clinical picture and virus shedding in these animals. To be able to analyse this, challenge experiments in relevant bat species that are available for such experiments are necessary. This was first carried out in R. aegyptiacus fruit bats using SARS-CoV isolated from R. sinicus bats (Ge et al. 2013, van Doremalen et al. 2018). In this study, R. aegyptiacus bats did not develop any clinical signs, displayed a mild involvement of the respiratory tract and only produced low levels of antibodies against the viral proteins. A challenge study using a human SARS-CoV-2 isolate in the same bat species again revealed results indicating a potential role of these bats as a reservoir host, since the animals also showed no clinical signs, but shed virus, transmitted the infection to contact animals and seroconverted (Schlottau et al. 2020). In a different study Eptesicus fuscus bats were experimentally challenged with SARS-CoV-2 (Hall et al. 2020): In contrast to R. aegyptiacus, E. fuscus bats seem to be resistant to infection with SARS-CoV-2. No virus was detectable in tissues and no viral excretion was observed, demonstrating that certain bat species serve as reservoirs for certain viruses. Such challenge experiments are indispensable for the assessment of the role of the infected bat species or closely related bat species in the epidemiology of the different bat CoVs. It must however be kept in mind, that bat species available for such experiments are not necessarily the most relevant species for a certain disease agent, which may compromise the interpretation of the results.
Conclusions and Outlook
Relevance for public health
CoVs infect a wide variety of animals and humans, causing diseases of the respiratory, enteric, hepatic and neurological systems with different severity (Wong et al. 2019). Because of their special mechanisms of viral replication with possible recombinations and mutation (Graham and Baric 2010, Lau et al. 2010, Tian et al. 2014), CoVs may adapt rapidly to new hosts upon spillover infections. On the other hand, with 1300-1400 species (American Society of Mammalogists 2020, Fenton 2015, Teeling et al. 2018), bats account for about 20% of all mammalian species (Teeling et al. 2018), and harbour a large number of viruses (Han et al. 2015). In bat colonies the physical proximity facilitates viral transmission among bats, supporting the virus circulation within the population (Han et al. 2015).
With more than 30 complete bat-CoV genomes sequenced within the last 17 years, bats are the group of mammals harbouring the largest number and variety of CoVs, and bat-CoVs may in fact be the ancestors for alpha- and beta-CoVs (Wong et al. 2019). Rhinolophus bats were shown to be natural reservoirs for SARS-like CoVs (Ge et al. 2013, Lau et al. 2005, Li et al. 2005) and several beta-CoVs detected in bats are closely related to MERS-CoV (Anthony et al. 2017a, Corman et al. 2014) and SARS-CoV-2 (Boni et al. 2020, Wacharapluesadee et al. 2021). Spill-over events of CoVs from bats to humans have led to devastating outbreaks, so intense surveillance of bats and their CoVs has tremendous relevance for public health. The worldwide distribution, roosting behaviour of bats close to or even in human settlements resulting in close contacts to humans may expose the public to a general risk of infection with bat-borne viruses (Han et al. 2015), such as bat-CoVs. The ability of bats to fly thousands of kilometres (Neuweiler 2000), may also contribute to long-distance dispersal of viruses (Mandl et al. 2018, Smith and Wang 2013). However, these findings should not be misinterpreted in a way to support programmes aimed at a decrease or removal of bat populations in the vicinity of human populations. Bats play a crucial role in the ecosystem (Kunz et al. 2011) and their protection should be a primary goal when implementing public health measures.
Research gaps
As a reaction to the SARS-CoV and lately the SARS-CoV-2 outbreaks, research on bats and CoVs has been intensified. A literature search shows that the SARS-CoV emergence resulted in the publication of approximately 4000 scientific articles within the first five years, while almost 11.000 articles related to SARS-CoV-2 have already been uploaded as preprint or peer-reviewed article within the last 10 months (bioRxiv 2020). This is a perfect example for event driven research that can obviously only react to outbreaks that have occurred in the past. Due to their high frequency of recombination and the large number of CoV species, a number of studies have highlighted the possible relevance of CoVs for future pandemic situations prior to the COVID19-pandemic, even predicting „disastrous consequences“ (Woo et al. 2006b), and considering CoVs a „time bomb“ (Cheng et al. 2007). To decrease the risk of future pandemic situations caused by CoVs or other disease agents, a preventive approach would be more beneficial, e.g. identifying general replication strategies of virus families or virus genera that are suspected to carry a pandemic risk, in order to develop intervention strategies that will at least provide us with initial measures to control a disease outbreak in its early stage and gain time to develop more targeted interventions. At the same time, vaccine strategies should be developed that can quickly and easily be adapted to any virus, so called “virus X”, that may emerge or re-emerge in the future. Such activities have been ongoing for the last years, and have now considerably accelerated the development of promising vaccine candidates against COVID-19.
Necessary measures and precautions
In order to get to a situation where we are able to act in a preventive manner instead of being forced to react to new outbreaks, we need to monitor the emergence of new viruses. Surveillance should be focused in regions and putative reservoir species that have been the origin of large disease outbreaks in the past. Of particular focus are tropical and subtropical regions (e.g. the Chinese Guangdong Wuhan regions as the putative regions of origin for SARS-CoV and SARS-CoV-2; African countries as the origin of Ebola disease outbreaks and South-East-Asian regions as regions, where outbreaks with highly pathogenic Nipah and Hendra viruses have occurred). All these viruses have been postulated to be transmitted directly or indirectly from bats to humans. Therefore, regular monitoring of bats and other potential reservoir hosts, especially, but not exclusively in tropical and subtropical regions, enables a science-based risk assessment. Factors like habitat destruction, human encroachment or climate change as potential drivers for viral spill-over events and disease emergence should be included in the evaluation. Given the high-throughput sequencing methods that are now available often as portable devices, such screening activities no longer need to be focused on specific disease agents, but can be performed in a more general, unbiased approach.
Acknowledgement
The authors are grateful to Franz Conraths and Matthew Pickin for their critical review and copy-editing of the manuscript.
Ethical approval
Not applicable.
Conflict of interest
The authors hereby declare that they have no proprietary, professional or other personal interests in any product, service and/or company that could have influenced the contents or opinions expressed in this publication.
Funding
Not applicable.
Authors contribution
Both authors drafted and approved the article with equal contributions.
Address for correspondence
Dr. Anne Balkema-Buschmann
Friedrich-Loeffler-Institut
Institute of Novel and Emerging Infectious Diseases
Südufer 10
17493 Greifswald – Insel Riems, Germany
Anne.Balkema-Buschmann@fli.de
References
American Society of Mammalogists (2020): Mammal Diversity Database. https://mammaldiversity.org (accessed 06.04.2021).
Annan A, Baldwin H, Corman VM, Klose S, Owusu M, Nkrumah EE, Badu EK, Anti P, Agbenyega O, Meyer B, Oppong S, Sarkodie YA, Kalko EKV, Lina PHC, Godlevska E, Reusken C, Seebens A, Gloza-Rausch F, Vallo P, Tschapka M, Drosten C, Drexler JF (2013): Human Betacoronavirus 2c EMC/2012–related Viruses in Bats, Ghana and Europe. Emerg Infect Dis 19: 456–459.
Anthony SJ, Ojeda-Flores R, Rico-Chávez O, Navarrete-Macias I, Zambrana-Torrelio CM, Rostal MK, Epstein JH, Tipps T, Liang E, Sanchez-Leon M, Sotomayor-Bonilla J, Aguirre AA, Ávila-Flores R, Medellín RA, Goldstein T, Suzán G, Daszak P, Lipkin WI (2013): Coronaviruses in bats from Mexico. J Gen Virol 94: 1028–1038.
Anthony SJ, Gilardi K, Menachery VD, Goldstein T, Ssebide B, Mbabazi R, Navarrete-Macias I, Liang E, Wells H,
Hicks A, Petrosov A, Byarugaba DK, Debbink K, Dinnon KH, Scobey T, Randell SH, Yount BL, Cranfield M,
Johnson CK, Baric RS, Lipkin WI, Mazet JAK (2017a): Further Evidence for Bats as the Evolutionary Source of Middle East Respiratory Syndrome Coronavirus. mBio 8: e00373–00317.
Anthony SJ, Johnson CK, Greig DJ, Kramer S, Che X,
Wells H, Hicks AL, Joly DO, Wolfe ND, Daszak P,
Karesh W, Lipkin WI, Morse SS, Mazet JAK, Goldstein T (2017b): Global patterns in coronavirus diversity. Virus Evol 3: vex012.
August TA, Mathews F, Nunn MA (2012): Alphacoronavirus Detected in Bats in the United Kingdom. Vector Borne Zoonotic Dis 12: 530–533.
Badrane H, Tordo N (2001): Host switching in Lyssavirus history from the Chiroptera to the Carnivora orders. J Virol 75: 8096–8104.
Balboni A, Palladini A, Bogliani G, Battilani M (2011): Detection of a virus related to betacoronaviruses in Italian greater horseshoe bats. Epidemiol Infect 139: 216–219.
Balboni A, Gallina L, Palladini A, Prosperi S, Battilani M (2012): A Real-Time PCR Assay for Bat SARS-Like Coronavirus Detection and Its Application to Italian Greater Horseshoe Bat Faecal Sample Surveys. ScientificWorldJournal 2012: 989514.
Banerjee A, Kulcsar K, Misra V, Frieman M, Mossman K (2019): Bats and Coronaviruses. Viruses 11: 41.
Banerjee A, Baker ML, Kulcsar K, Misra V, Plowright R, Mossman K (2020): Novel insights into immune systems of bats. Front Immunol 11: 26.
bioRxiv (2020): The preprint server for biology. https://www.biorxiv.org/ (accessed 15.11.2020).
Boardman WSJ, Baker ML, Boyd V, Crameri G, Peck GR, Reardon T, Smith IG, Caraguel CGB, Prowse TAA (2020): Serological evidence of exposure to a coronavirus antigenically related to severe acute respiratory syndrome virus (SARS-CoV-1) in the Grey-headed flying fox (Pteropus poliocephalus). Transbound Emerg Dis DOI: 10.1111/tbed.13908.
Boni MF, Lemey P, Jiang X, Lam TT, Perry BW, Castoe TA, Rambaut A, Robertson DL (2020): Evolutionary origins of the SARS-CoV-2 sarbecovirus lineage responsible for the COVID-19 pandemic. Nat Microbiol 5: 1408–1417.
Bonny TS, Driver JP, Paisie T, Salemi M, Morris JG, Shender LA, Smith L, Enloe C, Oxenrider K, Gore JA, Loeb JC, Wu CY, Lednicky JA (2017): Detection of Alphacoronavirus vRNA in the Feces of Brazilian Free-Tailed Bats (Tadarida brasiliensis) from a Colony in Florida, USA. Diseases 5: 7.
Bourgarel M, Pfukenyi DM, Boué V, Talignani L, Chiweshe N, Diop F, Caron A, Matope G, Missé D, Liégeois F (2018): Circulation of Alphacoronavirus, Betacoronavirus and Paramyxovirus in Hipposideros bat species in Zimbabwe. Infect Genet Evol 58: 253–257.
Bronrier GN, Maloney SK, Buffenstein R (1999): Survival tactics within thermally-challenging roosts: heat tolerance and cold sensitivity in the Angolan free-tailed bat, Mops condylurus. S Afr J Zool 34: 1–10.
Calisher CH, Childs JE, Field HE, Holmes KV, Schountz T (2006): Bats: important reservoir hosts of emerging viruses. Clin Microbiol Rev 19: 531–545.
Chen L, Liu B, Yang J, Jin Q (2014): DBatVir: the database of bat-associated viruses. Database (Oxford) 2014: bau021.
Cheng VC, Lau SK, Woo PC, Yuen KY (2007): Severe acute respiratory syndrome coronavirus as an agent of emerging and reemerging infection. Clin Microbiol Rev 20: 660–694.
Chu DKW, Poon LLM, Chan KH, Chen H, Guan Y, Yuen KY, Peiris JSM (2006): Coronaviruses in bent-winged bats (Miniopterus spp.). J Gen Virol 87: 2461–2466.
Corman VM, Rasche A, Diallo TD, Cottontail VM, Stöcker A, Souza B, Corrêa JI, Carneiro AJB, Franke CR, Nagy M, Metz M, Knörnschild M, Kalko EKV, Ghanem SJ, Morales KDS, Salsamendi E, Spínola M, Herrler G, Voigt CC, Tschapka M, Drosten C, Drexler JF (2013): Highly diversified coronaviruses in neotropical bats. J Gen Virol 94: 1984–1994.
Corman VM, Ithete NL, Richards LR, Schoeman MC, Preiser W, Drosten C, Drexler JF (2014): Rooting the Phylogenetic Tree of Middle East Respiratory Syndrome Coronavirus by Characterization of a Conspecific Virus from an African Bat. J Virol 88: 11297–11303.
Corman VM, Baldwin HJ, Tateno AF, Zerbinati RM, Annan A, Owusu M, Nkrumah EE, Maganga GD, Oppong S, Adu-Sarkodie Y, Vallo P, da Silva Filho LVRF, Leroy EM, Thiel V, van der Hoek L, Poon LLM, Tschapka M, Drosten C, Drexler JF (2015): Evidence for an Ancestral Association of Human Coronavirus 229E with Bats. J Virol 89: 11858–11870.
Corman VM, Muth D, Niemeyer D, Drosten C (2018): Hosts and Sources of Endemic Human Coronaviruses. Adv Virus Res 100: 163–188.
De Benedictis P, Marciano S, Scaravelli D, Priori P, Zecchin B, Capua I, Monne I, Cattoli G (2014): Alpha and lineage C betaCoV infections in Italian bats. Virus Genes 48: 366–371.
De Sabato L, Lelli D, Faccin F, Canziani S, Di Bartolo I, Vaccari G, Moreno A (2019): Full genome characterization of two novel Alpha-coronavirus species from Italian bats. Virus Res 260: 60–66.
Delmas O, Holmes EC, Talbi C, Larrous F, Dacheux L, Bouchier C, Bourhy H (2008): Genomic diversity and evolution of the lyssaviruses. PLoS One 3: e2057–e2057.
Dominguez SR, O’Shea TJ, Oko LM, Holmes KV (2007): Detection of group 1 coronaviruses in bats in North America. Emerg Infect Dis 13: 1295–1300.
Drexler JF, Gloza-Rausch F, Glende J, Corman VM, Muth D, Goettsche M, Seebens A, Niedrig M, Pfefferle S, Yordanov S, Zhelyazkov L, Hermanns U, Vallo P, Lukashev A, Müller MA, Deng H, Herrler G, Drosten C (2010): Genomic characterization of severe acute respiratory syndrome-related coronavirus in European bats and classification of coronaviruses based on partial RNA-dependent RNA polymerase gene sequences. J Virol 84: 11336–11349.
Drexler JF, Corman VM, Muller MA, Maganga GD, Vallo P, Binger T, Gloza-Rausch F, Cottontail VM, Rasche A, Yordanov S, Seebens A, Knornschild M, Oppong S, Adu Sarkodie Y, Pongombo C, Lukashev AN, Schmidt-Chanasit J, Stocker A, Carneiro AJ, Erbar S, Maisner A, Fronhoffs F, Buettner R, Kalko EK, Kruppa T, Franke CR, Kallies R, Yandoko ER, Herrler G, Reusken C, Hassanin A, Kruger DH, Matthee S, Ulrich RG, Leroy EM, Drosten C (2012): Bats host major mammalian paramyxoviruses. Nat Commun 3: 796.
Drexler JF, Corman VM, Drosten C (2014): Ecology, evolution and classification of bat coronaviruses in the aftermath of SARS. Antiviral Res 101: 45–56.
Drosten C, Günther S, Preiser W, van der Werf S, Brodt HR, Becker S, Rabenau H, Panning M, Kolesnikova L, Fouchier RA, Berger A, Burguière AM, Cinatl J, Eickmann M, Escriou N, Grywna K, Kramme S, Manuguerra JC, Müller S, Rickerts V, Stürmer M, Vieth S, Klenk HD, Osterhaus AD, Schmitz H, Doerr HW (2003): Identification of a novel coronavirus in patients with severe acute respiratory syndrome. N Engl J Med 348: 1967–1976.
Falcón A, Vázquez-Morón S, Casas I, Aznar C, Ruiz G, Pozo F, Perez-Breña P, Juste J, Ibáñez C, Garin I, Aihartza J, Echevarría JE (2011): Detection of alpha and betacoronaviruses in multiple Iberian bat species. Arch Virol 156: 1883–1890.
Fan Y, Zhao K, Shi ZL, Zhou P (2018): Bat Coronaviruses in China. Viruses 11(3): 210. DOI: 10.3390/v11030210.
Fenton MB (2015): Bats: A World of Science and Mystery. University of Chicago Press, Chicago, USA.
Fischer K, Zeus V, Kwasnitschka L, Kerth G, Haase M, Groschup MH, Balkema-Buschmann A (2016): Insectivorous bats carry host specific astroviruses and coronaviruses across different regions in Germany. Infect Genet Evol 37: 108–116.
Forbes KM, Webala PW, Jaaskelainen AJ, Abdurahman S, Ogola J, Masika MM, Kivisto I, Alburkat H, Plyusnin I, Levanov L, Korhonen EM, Huhtamo E, Mwaengo D, Smura T, Mirazimi A, Anzala O, Vapalahti O, Sironen T (2019): Bombali Virus in Mops condylurus Bat, Kenya. Emerg Infect Dis Emerg Infect Dis 25: 955–957.
Ge XY, Li JL, Yang XL, Chmura AA, Zhu G, Epstein JH, Mazet JK, Hu B, Zhang W, Peng C, Zhang YJ, Luo CM, Tan B, Wang N, Zhu Y, Crameri G, Zhang SY, Wang LF, Daszak P, Shi ZL (2013): Isolation and characterization of a bat SARS-like coronavirus that uses the ACE2 receptor. Nature 503: 535–538.
Geldenhuys M, Weyer J, JW, Nel LH, Markotter W (2013): Coronaviruses in South African bats. Vector Borne Zoonotic Dis 13:516–519.
Gloza-Rausch F, Ipsen A, Seebens A, Göttsche M, Panning M, Bispo de Filippis AM, Petersen N, Annan A, Grywna K, Müller M, Pfefferle S, Park SS (2008): Detection and Prevalence Patterns of Group I Coronaviruses in Bats, Northern Germany. Emerg Infect Dis 14: 626–631.
Goldstein T, Anthony SJ, Gbakima A, Bird BH, Bangura J, Tremeau-Bravard A, Belaganahalli MN, Wells HL, Dhanota JK, Liang E, Grodus M, Jangra RK, DeJesus VA, Lasso G, Smith BR, Jambai A, Kamara BO, Kamara S, Bangura W, Monagin C, Shapira S, Johnson CK, Saylors K, Rubin EM, Chandran K, Lipkin WI, Mazet JAK (2018): The discovery of Bombali virus adds further support for bats as hosts of ebolaviruses. Nat Microbiol 3: 1084–1089.
Gong L, Li J, Zhou Q, Xu Z, Chen L, Zhang Y, Xue C, Wen Z, Cao Y (2017): A New Bat-HKU2-like Coronavirus in Swine, China, 2017. Emerg Infect Dis 23: 1607–1609.
Gorbunova V, Seluanov A, Kennedy BK (2020): The World Goes Bats: Living Longer and Tolerating Viruses. Cell Metab 32: 31–43.
Gouilh MA, Puechmaille SJ, Gonzalez JP, Teeling E, Kittayapong P, Manuguerra JC (2011): SARS-Coronavirus ancestor’s foot-prints in South-East Asian bat colonies and the refuge theory. Infect Genet Evol 11: 1690–1702.
Graham RL, Baric RS (2010): Recombination, Reservoirs, and the Modular Spike: Mechanisms of Coronavirus Cross-Species Transmission. J Virol 84: 3134–3146.
Guan Y, Zheng BJ, He YQ, Liu XL, Zhuang ZX, Cheung CL, Luo SW, Li PH, Zhang LJ, Guan YJ, Butt KM, Wong KL, Chan KW, Lim W, Shortridge KF, Yuen KY, Peiris JS, Poon LL (2003): Isolation and characterization of viruses related to the SARS coronavirus from animals in southern China. Science 302: 276–278.
Guo Y-R, Cao Q-D, Hong Z-S, Tan Y-Y, Chen S-D, Jin H-J,
Tan K-S, Wang D-Y, Yan Y (2020): The origin, transmission and clinical therapies on coronavirus disease 2019
(COVID-19) outbreak – an update on the status. Mil Med Res 7: 11.
Hall JS, Knowles S, Nashold SW, Ip HS, Leon AE, Rocke T, Keller S, Carossino M, Balasuriya U, Hofmeister E (2020): Experimental challenge of a North American bat species, big brown bat (Eptesicus fuscus), with SARS-CoV-2. Transbound Emerg Dis. DOI: 10.1111/tbed.13949.
Han HJ, Wen HL, Zhou CM, Chen FF, Luo LM, Liu JW, Yu XJ (2015): Bats as reservoirs of severe emerging infectious diseases. Virus Res 205: 1–6.
Hayman DTS (2016): Bats as Viral Reservoirs. Annual Review of Virology 3(1): 77-99.
He X, Lau EHY, Wu P, Deng X, Wang J, Hao X, Lau YC,
Wong JY, Guan Y, Tan X, Mo X, Chen Y, Liao B, Chen W, Hu F, Zhang Q, Zhong M, Wu Y, Zhao L, Zhang F, Cowling BJ, Li F, Leung GM (2020): Temporal dynamics in viral
shedding and transmissibility of COVID-19. Nat Med 26: 672–675.
Hu B, Ge X, Wang LF, Shi Z (2015): Bat origin of human coronaviruses. Virol J 12: 221–221.
Huynh J, Li S, Yount B, Smith A, Sturges L, Olsen JC, Nagel J, Johnson JB, Agnihothram S, Gates JE, Frieman MB, Baric RS, Donaldson EF (2012): Evidence supporting a zoonotic origin of human coronavirus strain NL63. J Virol 86: 12816–12825.
Irving AT, Ahn M, Goh G, Anderson DE, Wang LF (2021): Lessons from the host defences of bats, a unique viral reservoir. Nature 589: 363–370.
Ithete NL, Stoffberg S, Corman VM, Cottontail V, Richards LR, Schoeman MC, Drosten C, Drexler JF, Preiser W (2013): Close Relative of Human Middle East Respiratory Syndrome Coronavirus in Bat, South Africa. Emerg Infect Dis 19:1697–1699.
Joffrin L, Goodman SM, Wilkinson DA, Ramasindrazana B, Lagadec E, Gomard Y, Le Minter G, Dos Santos A, Corrie Schoeman M, Sookhareea R, Tortosa P, Julienne S, Gudo ES, Mavingui P, Lebarbenchon C (2020): Bat coronavirus phylogeography in the Western Indian Ocean. Sci Rep 10: 6873.
Johns Hopkins University – Coronavirus Resource Center (2020): Coronavirus COVID-19 Global Cases by the Center for Systems Science and Engineering (CSSE) at Johns Hopkins University (JHU). https://coronavirus.jhu.edu/map.html (accessed 02.04.2021).
Jones G, Teeling EC (2006): The evolution of echolocation in bats. Trends Ecol Evol 21: 149–156.
Karan LS, Makenov MT, Korneev MG, Sacko N, Boumbaly S, Yakovlev SA, Kourouma K, Bayandin RB, Gladysheva AV, Shipovalov AV, Yurganova IA, Grigorieva YE, Fedorova MV, Scherbakova SA, Kutyrev VV, Agafonov AP, Maksyutov RA, Shipulin GA, Maleev VV, Boiro M, Akimkin VG, Popova AY (2019): Bombali Virus in Mops condylurus Bats, Guinea. Emerg Infect Dis 25:1774–1775.
Kemenesi G, Dallos B, Görföl T, Boldogh S, Estók P, Kurucz K, Kutas A, Földes F, Oldal M, Németh V, Martella V, Bányai K, Jakab F (2014): Molecular Survey of RNA Viruses in Hungarian Bats: Discovering Novel Astroviruses, Coronaviruses, and Caliciviruses. Vector Borne Zoonotic Dis 14: 846–855.
Kemenesi G, Zhang D, Marton S, Dallos B, Görföl T, Estók P, Boldogh S, Kurucz K, Oldal M, Kutas A, Bányai K, Jakab F (2015): Genetic characterization of a novel picornavirus detected in Miniopterus schreibersii bats. J Gen Virol 96: 815–821.
Killerby ME, Biggs HM, Midgley CM, Gerber SI, Watson JT (2020): Middle East Respiratory Syndrome Coronavirus Transmission. Emerg Infect Dis 26:191–198.
Kunz TH, Braun de Torrez E, Bauer D, Lobova T, Fleming TH (2011): Ecosystem services provided by bats. Ann N Y Acad Sci 1223: 1–38.
Lacroix A, Vidal N, Keita AK, Thaurignac G, Esteban A, De Nys H, Diallo R, Toure A, Goumou S, Soumah AK, Povogui M, Koivogui J, Monemou J-L, Raulino R, Nkuba A, Foulongne V, Delaporte E, Ayouba A, Peeters M (2020): Wide Diversity of Coronaviruses in Frugivorous and Insectivorous Bat Species: A Pilot Study in Guinea, West Africa. Viruses 12: 855.
Lam TT, Jia N, Zhang YW, Shum MH, Jiang JF, Zhu HC, Tong YG, Shi YX, Ni XB, Liao YS, Li WJ, Jiang BG, Wei W, Yuan TT, Zheng K, Cui XM, Li J, Pei GQ, Qiang X, Cheung WY, Li LF, Sun FF, Qin S, Huang JC, Leung GM, Holmes EC, Hu YL, Guan Y, Cao WC (2020): Identifying SARS-CoV-2-related coronaviruses in Malayan pangolins. Nature 583: 282–285.
Lau SK, Woo PC, Li KS, Huang Y, Tsoi HW, Wong BH, Wong SS, Leung SY, Chan KH, Yuen KY (2005): Severe acute respiratory syndrome coronavirus-like virus in Chinese horseshoe bats. Proc Natl Acad Sci U S A 102: 14040–14045.
Lau SK, Woo PC, Li KS, Huang Y, Wang M, Lam CS, Xu H, Guo R, Chan KH, Zheng BJ, Yuen KY (2007): Complete genome sequence of bat coronavirus HKU2 from Chinese horseshoe bats revealed a much smaller spike gene with a different evolutionary lineage from the rest of the genome. Virology 367: 428–439.
Lau SK, Li KSM, Huang Y, Shek C-T, Tse H, Wang M, Choi GKY, Xu H, Lam CSF, Guo R, Chan K-H, Zheng B-J, Woo PCY, Yuen K-Y (2010): Ecoepidemiology and complete genome comparison of different strains of Severe Acute Respiratory Syndrome-Related Rhinolophus Bat Coronavirus in China reveal bats as a reservoir for acute, self-limiting infection that allows recombination events. J Virol 84: 2808–2819.
Lau SK, Wong ACP, Lau TCK, Woo PCY (2017): Molecular Evolution of MERS Coronavirus: Dromedaries as a Recent Intermediate Host or Long-Time Animal Reservoir? Int J Mol Sci 18: 2138.
Lazov CM, Chriél M, Baagøe HJ, Fjederholt E, Deng Y, Kooi EA, Belsham GJ, Bøtner A, Rasmussen TB (2018): Detection and Characterization of Distinct Alphacoronaviruses in Five Different Bat Species in Denmark. Viruses 10: 486.
Le Poder S (2011): Feline and canine coronaviruses: common genetic and pathobiological features. Adv Virol 2011: 609465.
Lecis R, Mucedda M, Pidinchedda E, Pittau M, Alberti A (2019): Molecular identification of Betacoronavirus in bats from Sardinia (Italy): first detection and phylogeny. Virus Genes 55: 60–67.
Lelli D, Papetti A, Sabelli C, Rosti E, Moreno A, Boniotti MB (2013): Detection of coronaviruses in bats of various species in Italy. Viruses 5: 2679–2689.
Letko M, Seifert SN, Olival KJ, Plowright RK, Munster VJ (2020): Bat-borne virus diversity, spillover and emergence. Nat Rev Microbiol 18: 461–471.
Li W, Shi Z, Yu M, Ren W, Smith C, Epstein JH, Wang H, Crameri G, Hu Z, Zhang H, Zhang J, McEachern J, Field H, Daszak P, Eaton BT, Zhang S, Wang LF (2005): Bats are natural reservoirs of SARS-like coronaviruses. Science 310: 676–679.
Lima FE, Campos FS, Kunert Filho HC, Batista HB, Carnielli P, Jr., Cibulski SP, Spilki FR, Roehe PM, Franco AC (2013): Detection of Alphacoronavirus in velvety free-tailed bats (Molossus molossus) and Brazilian free-tailed bats (Tadarida brasiliensis) from urban area of Southern Brazil. Virus Genes 47: 164–167.
Luo CM, Wang N, Yang XL, Liu HZ, Zhang W, Li B, Hu B, Peng C, Geng QB, Zhu GJ, Li F, Shi ZL (2018): Discovery of novel bat coronaviruses in South China that use the same receptor as Middle East Respiratory Syndrome Coronavirus. J Virol 92: e00116-18.
Maganga GD, Pinto A, Mombo IM, Madjitobaye M, Mbeang Beyeme AM, Boundenga L, Ar Gouilh M, N’Dilimabaka N, Drexler JF, Drosten C, Leroy EM (2020): Genetic diversity and ecology of coronaviruses hosted by cave-dwelling bats in Gabon. Sci Rep 10: 7314–7314.
Maloney SK, Bronner GN, Buffenstein R (1999): Thermoregulation in the Angolan free-tailed bat Mops condylurus: A small mammal that uses hot roosts. Physiol Biochem Zool 72: 385–396.
Mandl JN, Schneider C, Schneider DS, Baker ML (2018): Going to bat(s) for studies of disease tolerance. Front Immunol 9: 2112–2112.
Markotter W, Geldenhuys M, Jansen van Vuren P, Kemp A, Mortlock M, Mudakikwa A, Nel L, Nziza J, Paweska J, Weyer J (2019): Paramyxo- and Coronaviruses in Rwandan Bats. Trop Med Infect Dis 4: 99.
Misra V, Dumonceaux T, Dubois J, Willis C, Nadin-Davis S, Severini A, Wandeler A, Lindsay R, Artsob H (2009): Detection of polyoma and corona viruses in bats of Canada. J Gen Virol 90: 2015–2022.
Mollentze N, Streicker DG (2020): Viral zoonotic risk is homogenous among taxonomic orders of mammalian and avian reservoir hosts. Proc Natl Acad Sci U S A 117: 9423–9430.
Monchatre-Leroy E, Boué F, Boucher J-M, Renault C, Moutou F, Ar Gouilh M, Umhang G (2017): Identification of Alpha and Beta Coronavirus in Wildlife Species in France: Bats, Rodents, Rabbits, and Hedgehogs. Viruses 9: 364.
Moratelli R, Calisher CH (2015): Bats and zoonotic viruses: can we confidently link bats with emerging deadly viruses? Mem Inst Oswaldo Cruz 110: 1–22.
Moreno A, Lelli D, de Sabato L, Zaccaria G, Boni A, Sozzi E, Prosperi A, Lavazza A, Cella E, Castrucci MR, Ciccozzi M, Vaccari G (2017): Detection and full genome characterization of two beta CoV viruses related to Middle East respiratory syndrome from bats in Italy. Virol J 14: 239–239.
Müller MA, Paweska JT, Leman PA, Drosten C, Grywna K, Kemp A, Braack L, Sonnenberg K, Niedrig M, Swanepoel R (2007): Coronavirus antibodies in African bat species. Emerg Infect Dis 13: 1367–1370.
Nabi G, Wang Y, Lü L, Jiang C, Ahmad S, Wu Y, Li D (2021): Bats and birds as viral reservoirs: A physiological and ecological perspective. Sci Total Environ 754:142372.
Neuweiler G (2000): The Biology of Bats. Oxford University Press, New York.
Nziza J, Goldstein T, Cranfield M, Webala P, Nsengimana O, Nyatanyi T, Mudakikwa A, Tremeau-Bravard A, Byarugaba D, Tumushime JC, Mwikarago IE, Gafarasi I, Mazet J, Gilardi K (2019): Coronaviruses Detected in Bats in Close Contact with Humans in Rwanda. EcoHealth 16: 1–3.
O’Shea TJ, Cryan PM, Cunningham AA, Fooks AR, Hayman DT, Luis AD, Peel AJ, Plowright RK, Wood JL (2014): Bat flight and zoonotic viruses. Emerg Infect Dis 20: 741–745.
Osborne C, Cryan PM, O’Shea TJ, Oko LM, Ndaluka C, Calisher CH, Berglund AD, Klavetter ML, Bowen RA, Holmes KV, Dominguez SR (2011): Alphacoronaviruses in New World bats: prevalence, persistence, phylogeny, and potential for interaction with humans. PLoS One 6: e19156–e19156.
Paden CR, Yusof M, Al Hammadi ZM, Queen K, Tao Y, Eltahir YM, Elsayed EA, Marzoug BA, Bensalah OKA, Khalafalla AI, Al Mulla M, Khudhair A, Elkheir KA, Issa ZB, Pradeep K, Elsaleh FN, Imambaccus H, Sasse J, Weber S, Shi M, Zhang J, Li Y, Pham H, Kim L, Hall AJ, Gerber SI, Al Hosani FI, Tong S, Al Muhairi SSM (2018): Zoonotic origin and transmission of Middle East respiratory syndrome coronavirus in the UAE. Zoonoses Public Health 65: 322–333.
Pauly M, Pir JB, Loesch C, Sausy A, Snoeck CJ, Hübschen JM, Muller CP (2017): Novel Alphacoronaviruses and Paramyxoviruses Cocirculate with Type 1 and Severe Acute Appl Environ Microbiol 83: e01326-17.
Peiris JS, Yuen KY, Osterhaus AD, Stöhr K (2003): The severe acute respiratory syndrome. N Engl J Med 349: 2431–2441.
Pfefferle S, Oppong S, Bispo de Filippis AM, Gloza-Rausch F, Ipsen A, Seebens A, Müller MA, Annan A, Vallo P, Adu-Sarkodie Y, Kruppa TF, Park SS (2009): Distant Relatives of Severe Acute Respiratory Syndrome Coronavirus and Close Relatives of Human Coronavirus 229E in Bats, Ghana. Emerg Infect Dis 15: 1377.
Plowright RK, Eby P, Hudson PJ, Smith IL, Westcott D, Bryden WL, Middleton D, Reid PA, McFarlane RA, Martin G, Tabor GM, Skerratt LF, Anderson DL, Crameri G, Quammen D, Jordan D, Freeman P, Wang LF, Epstein JH, Marsh GA, Kung NY, McCallum H (2015): Ecological dynamics of emerging bat virus spillover. Proc Biol Sci 282: 20142124.
Plowright RK, Parrish CR, McCallum H, Hudson PJ, Ko AI, Graham AL, Lloyd-Smith JO (2017): Pathways to zoonotic spillover. Nat Rev Microbiol 15: 502–510.
Poon LL, Chu DK, Chan KH, Wong OK, Ellis TM, Leung YH, Lau SK, Woo PC, Suen KY, Yuen KY, Guan Y, Peiris JS (2005): Identification of a novel coronavirus in bats. J Virol 79: 2001–2009.
Quan P-L, Firth C, Street C, Henriquez JA, Petrosov A, Tashmukhamedova A, Hutchison SK, Egholm M, Osinubi MOV, Niezgoda M, Ogunkoya AB, Briese T, Rupprecht CE, Lipkin WI (2010): Identification of a Severe Acute Respiratory Syndrome Coronavirus-Like Virus in a Leaf-Nosed Bat in Nigeria. mBio 1: e00208-00210.
Razanajatovo NH, Nomenjanahary LA, Wilkinson DA, Razafimanahaka JH, Goodman SM, Jenkins RK, Jones JPG, Heraud J-M (2015): Detection of new genetic variants of Betacoronaviruses in Endemic Frugivorous Bats of Madagascar. Virol J 12: 42–42.
Reusken CBEM, Lina PHC, Pielaat A, Vries Ad, Dam-Deisz C, Adema J, Drexler JF, Drosten C, Kooi EA (2010): Circulation of Group 2 Coronaviruses in a bat species common to urban areas in Western Europe. Vector Borne Zoonotic Dis 10: 785–791.
Rihtarič D, Hostnik P, Steyer A, Grom J, Toplak I (2010): Identification of SARS-like coronaviruses in horseshoe bats (Rhinolophus hipposideros) in Slovenia. Arch Virol 155: 507–514.
Schlottau K, Rissmann M, Graaf A, Schön J, Sehl J, Wylezich C, Höper D, Mettenleiter TC, Balkema-Buschmann A, Harder T, Grund C, Hoffmann D, Breithaupt A, Beer M (2020): SARS-CoV-2 in fruit bats, ferrets, pigs, and chickens: an experimental transmission study. Lancet Microbe 1: e218–e225.
Shi Z (2013): Emerging infectious diseases associated with bat viruses. Sci China Life Sci 56: 678–682.
Smith CS, de Jong CE, Meers J, Henning J, Wang L, Field HE (2016): Coronavirus Infection and Diversity in Bats in the Australasian Region. EcoHealth 13: 72–82.
Smith I, Wang LF (2013): Bats and their virome: an important source of emerging viruses capable of infecting humans. Curr Opin Virol 3: 84–91.
Steyer A, Gutiérrez-Aguire I, Kolenc M, Koren S, Kutnjak D, Pokorn M, Poljšak-Prijatelj M, Racki N, Ravnikar M,
Sagadin M, Fratnik Steyer A, Toplak N (2013): High similarity of novel orthoreovirus detected in a child hospitalized with acute gastroenteritis to mammalian orthoreoviruses found in bats in Europe. J Clin Microbiol 51: 3818–3825.
Su S, Wong G, Shi W, Liu J, Lai ACK, Zhou J, Liu W, Bi Y, Gao GF (2016): Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses. Trends Microbiol 24: 490–502.
Subudhi S, Rapin N, Bollinger TK, Hill JE, Donaldson ME, Davy CM, Warnecke L, Turner JM, Kyle CJ, Willis CKR, Misra V (2017): A persistently infecting coronavirus in hibernating Myotis lucifugus, the North American little brown bat. J Gen Virol 98: 2297–2309.
Subudhi S, Rapin N, Misra V (2019): Immune system modulation and viral persistence in bats: Understanding viral spillover. Viruses 11: 192.
Tang XC, Zhang JX, Zhang SY, Wang P, Fan XH, Li LF, Li G, Dong BQ, Liu W, Cheung CL, Xu KM, Song WJ, Vijaykrishna D, Poon LL, Peiris JS, Smith GJ, Chen H, Guan Y (2006): Prevalence and genetic diversity of coronaviruses in bats from China. J Virol 80: 7481–7490.
Tao Y, Tang K, Shi M, Conrardy C, Li KSM, Lau SKP, Anderson LJ, Tong S (2012): Genomic characterization of seven distinct bat coronaviruses in Kenya. Virus Res 167: 67–73.
Tao Y, Shi M, Chommanard C, Queen K, Zhang J, Markotter W, Kuzmin IV, Holmes EC, Tong S (2017): Surveillance of bat coronaviruses in Kenya identifies relatives of human coronaviruses NL63 and 229E and their recombination history. J Virol 91: e01953-16.
Teeling E, Vernes S, Davalos L, Ray D, Gilbert M, Myers E, Santos T, Dool S, Tavares V, Lopes G, Power M, Botto G, Lim L-S, Consortium BK (2018): Bat biology, genomes, and the Bat1K Project: To generate chromosome-level genomes for all living bat species. Annu Rev Anim Biosci 15: 23–46.
Thomas DW (1983): The annual migrations of three species of West African fruit bats (Chiroptera: Pteropodidae). Can J Zool 61: 2266–2272.
Tian P-F, Jin Y-L, Xing G, Qv L-L, Huang Y-W, Zhou J-Y (2014): Evidence of recombinant strains of Porcine Epidemic Diarrhea Virus, United States, 2013. Emerg Infect Dis 20: 1731.
Tong S, Conrardy C, Ruone S, Kuzmin IV, Guo X, Tao Y, Niezgoda M, Haynes L, Agwanda B, Breiman RF, Anderson LJ, Rupprecht CE (2009): Detection of novel SARS-like and other coronaviruses in bats from Kenya. Emerg Infect Dis 15: 482.
van Doremalen N, Schäfer A, Menachery VD, Letko M, Bushmaker T, Fischer RJ, Figueroa DM, Hanley PW, Saturday G, Baric RS, Munster VJ (2018): SARS-like coronavirus WIV1-CoV does not replicate in Egyptian Fruit Bats (Rousettus aegyptiacus). Viruses 10: 727.
Vijaykrishna D, Smith GJ, Zhang JX, Peiris JS, Chen H, Guan Y (2007): Evolutionary insights into the ecology of coronaviruses. J Virol 81: 4012–4020.
Wacharapluesadee S, Tan CW, Maneeorn P, Duengkae P, Zhu F, Joyjinda Y, Kaewpom T, Chia WN, Ampoot W, Lim BL, Worachotsueptrakun K, Chen VC, Sirichan N, Ruchisrisarod C, Rodpan A, Noradechanon K, Phaichana T, Jantarat N, Thongnumchaima B, Tu C, Crameri G, Stokes MM, Hemachudha T, Wang LF (2021): Evidence for SARS-CoV-2 related coronaviruses circulating in bats and pangolins in Southeast Asia. Nat Commun 12: 972.
Waruhiu C, Ommeh S, Obanda V, Agwanda B, Gakuya F, Ge X-Y, Yang X-L, Wu L-J, Zohaib A, Hu B, Shi Z-L (2017): Molecular detection of viruses in Kenyan bats and discovery of novel astroviruses, caliciviruses and rotaviruses. Virol Sin 32: 101–114.
Wernery U, Lau SKP, Woo PCY (2017): Middle East respiratory syndrome (MERS) coronavirus and dromedaries. Vet J 220: 75–79.
Wong ACP, Li X, Lau SKP, Woo PCY (2019): Global epidemiology of bat coronaviruses. Viruses 11: 174.
Woo PCY, Lau SKP, Li KSM, Poon RWS, Wong BHL, Tsoi H-w, Yip BCK, Huang Y, Chan K-h, Yuen K-Y (2006a): Molecular diversity of coronaviruses in bats. Virology 351: 180–187.
Woo PCY, Lau SKP, Yip CCY, Huang Y, Tsoi H-W, Chan K-H, Yuen K-Y (2006b): Comparative analysis of 22 coronavirus HKU1 genomes reveals a novel genotype and evidence of natural recombination in coronavirus HKU1. J Virol 80: 7136–7145.
Woo PCY, Lau SKP, Lam CSF, Lau CCY, Tsang AKL, Lau JHN, Bai R, Teng JLL, Tsang CCC, Wang M, Zheng B-J, Chan K-H, Yuen K-Y (2012): Discovery of seven novel mammalian and avian coronaviruses in the genus deltacoronavirus supports bat coronaviruses as the gene source of alphacoronavirus and betacoronavirus and avian coronaviruses as the gene source of gammacoronavirus and deltacoronavirus. J Virol 86: 3995–4008.
World Health Organization (2019): Middle East respiratory syndrome coronavirus (MERS-CoV). https://www.who.int/news-room/fact-sheets/detail/middle-east-respirator…-(mers-cov) (accessed 02.04.2021).
World Health Organization (2020a): Timeline: WHO’s COVID-19 response. https://www.who.int/emergencies/diseases/novel-coronavirus-2019/interac… (accessed 07.04.2021).
World Health Organization (2020b): WHO Director-General’s opening remarks at the media briefing on COVID-19 - 11 March 2020. https://www.who.int/director-general/speeches/detail/who-director-gener… (accessed 07.04.2021).
Xu L, Zhang F, Yang W, Jiang T, Lu G, He B, Li X, Hu T, Chen G, Feng Y, Zhang Y, Fan Q, Feng J, Zhang H, Tu C (2016): Detection and characterization of diverse alpha- and betacoronavi–ruses from bats in China. Virol Sin 31(1): 69-77. DOI: 10.1007/s12250-016-3727-3.
Yang XL, Hu B, Wang B, Wang MN, Zhang Q, Zhang W, Wu LJ, Ge XY, Zhang YZ, Daszak P, Wang LF, Shi ZL (2015): Isolation and characterization of a novel bat coronavirus closely related to the direct progenitor of Severe Acute Respiratory Syndrome Coronavirus. J Virol 90: 3253–3256.
Ye ZW, Yuan S, Yuen KS, Fung SY, Chan CP, Jin DY (2020): Zoonotic origins of human coronaviruses. Int J Biol Sci 16: 1686–1697.
Zaki AM, van Boheemen S, Bestebroer TM, Osterhaus AD, Fouchier RA (2012): Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N Engl J Med 367: 1814–1820.
Zhou P, Fan H, Lan T, Yang XL, Shi WF, Zhang W, Zhu Y, Zhang YW, Xie QM, Mani S, Zheng XS, Li B, Li JM, Guo H, Pei GQ, An XP, Chen JW, Zhou L, Mai KJ, Wu ZX, Li D, Anderson DE, Zhang LB, Li SY, Mi ZQ, He TT, Cong F, Guo PJ, Huang R, Luo Y, Liu XL, Chen J, Huang Y, Sun Q, Zhang XL, Wang YY, Xing SZ, Chen YS, Sun Y, Li J, Daszak P, Wang LF, Shi ZL, Tong YG, Ma JY (2018): Fatal swine acute diarrhoea syndrome caused by an HKU2-related coronavirus of bat origin. Nature 556: 255–258.
Zhou P, Yang X-L, Wang X-G, Hu B, Zhang L, Zhang W, Si H-R, Zhu Y, Li B, Huang C-L, Chen H-D, Chen J, Luo Y, Guo H, Jiang R-D, Liu M-Q, Chen Y, Shen X-R, Wang X, Zheng X-S, Zhao K, Chen Q-J, Deng F, Liu L-L, Yan B, Zhan F-X, Wang Y-Y, Xiao G-F, Shi Z-L (2020): A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 579: 270–273.
Zhu N, Zhang D, Wang W, Li X, Yang B, Song J, Zhao X, Huang B, Shi W, Lu R, Niu P, Zhan F, Ma X, Wang D, Xu W, Wu G, Gao GF, Tan W (2020): A novel coronavirus from patients with pneumonia in China, 2019. N Engl J Med 382: 727–733.
Zumla A, Hui DS, Perlman S (2015): Middle East respiratory syndrome. Lancet 386: 995–1007.


Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2020-44-Balkema-Buschmann.pdf (0.26 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23761439-0299-2020-44-Balkema-Buschmann-Tabelle1.pdf (0.06 MB) herunterladen möchten

{kind=link}
{kind=link}