

Kastration von adulten männlichen C57BL/6JRj Mäusen ermöglicht Vergesellschaftung und Gruppenhaltung von zuvor einzeln gehaltenen Männchen: eine Harm-Benefit-Analyse
Berliner und Münchener Tierärztliche Wochenschrift 133
DOI: 10.2376/0005-9366-19055
© Schlütersche Verlagsgesellschaft mbH & Co. KG. 2020
Publiziert: 04/2020
Summary
Veterinary ethics play a critical role in animal research since veterinarians practicing in laboratory animal facilities face ethical dilemmas every day, such as the animal welfare issues that come along with standard laboratory housing. It is a common code of practice to keep mice in single-sex groups, although housing males together can result in stress or fatal injuries due to aggressive territorial and resource-related behaviour. However, if they are separated, social isolation also reduces their welfare. To eliminate aggression, 19 adult male C57BL/6JRj mice, which had been individually housed for up to 32 weeks, were castrated and resocialized in groups in this case report. Surgical intervention increased Mouse Grimace Scale scores for up to 150 min post-castration and concentrations of faecal corticosterone metabolites on day 2 post-castration when compared to baseline but did not affect burrowing and nesting, suggesting short-term stress and pain. Mice gained body weight within the following two weeks. After mice were resocialized in groups, a transient loss in body weight followed by weight gain reflected the process of habituation. The lack of fight-associated lesions and the high nest building performance provided evidence that the new groups were stable. Our investigation demonstrates that harmonious group-housing of castrated adult male C57BL/6JRj mice is possible. In terms of a lifetime harm-benefit analysis for the individual animal in this case report, the short-term post-surgical stress and pain may outweigh the long-lasting negative effects of single housing. Scientific data on the well-being of the mice provided the basis of the harm-benefit analysis.
Zusammenfassung
Die tiermedizinische Ethik spielt eine wichtige Rolle in der tierexperimentellen Forschung, da Tierärztinnen und Tierärzte in Versuchstierhaltungen täglich mit ethischen Dilemmata konfrontiert werden. Dazu zählen auch die tierschutzrelevanten Probleme, die auf die standardisierte Haltung der Versuchstiere zurückzuführen sind. Eine gängige Praxis ist die Haltung von Mäusen in gleichgeschlechtlichen Gruppen, obwohl Gruppenhaltung männlicher Mäuse aufgrund von aggressivem Territorialverhalten und Ressourcenkonflikten zu Stress bis hin zu lebensbedrohlichen Verletzungen führen kann. Werden sie separiert, beeinträchtigt die soziale Isolation ihr Wohlbefinden jedoch ebenfalls. Um das aggressive Verhalten zu reduzieren, wurden in diesem Fallbericht 19 adulte männliche C57BL/6JRj Mäuse, die bis zu 32 Wochen einzeln gehalten wurden, kastriert und in Gruppen resozialisiert. Der chirurgische Eingriff erhöhte die Mouse Grimace Scale Scores bis zu 150 Minuten und die Konzentration der fäkalen Kortikosteronmetabolite am Tag 2 nach Kastration im Vergleich zur Baseline, beeinflusste aber Wühl- und Nestbauverhalten nicht, was auf kurzfristigen Stress und Schmerz hindeutete. Die Mäuse nahmen innerhalb der darauffolgenden zwei Wochen an Körpergewicht zu. Nachdem die Tiere in Gruppen resozialisiert wurden, spiegelte sich die Habituation in einem vorübergehenden Verlust, gefolgt von einer Zunahme an Körpergewicht wider. Die Abwesenheit kampfassoziierter Läsionen und die hohe Nestbau-Performance indizierten, dass die neuen Gruppen stabil waren. Unsere Untersuchung zeigt, dass eine harmonische Gruppenhaltung von kastrierten adulten männlichen C57BL/6JRj Mäusen möglich ist. Hinsichtlich der Harm-Benefit-Analyse, bezogen auf das Gesamtleben eines Einzeltieres dieses Fallberichts, können die vorübergehenden postoperativen Schmerzen sowie Stress im Vergleich zu den langfristigen, negativen Effekten der Einzelhaltung toleriert werden. Die wissenschaftlich ermittelten Daten zum Wohlbefinden der Mäuse dienten als Basis der Harm-Benefit-Analyse.
Introduction
Laboratory animal science has always been an important field in veterinary ethics (Grimm et al. 2018). Procedures on animals causing pain, suffering, and distress raise difficult ethical and moral issues. Therefore, in a harm-benefit analysis, the benefits of the outcome are weighed against the harm the animal will experience (Grimm 2014, Grimm et al. 2019). This is required by law for animal experimentation (Directive 2010/63/EU), but is also applicable to housing, husbandry, and routine procedures diminishing the well-being of the animal. Veterinarians play a crucial role in this process because their expertise allows them to thoroughly evaluate the well-being of an animal. Accordingly, in the present case report, scientifically valid information obtained by veterinarians provided the basis of the harm-benefit analysis concerning a central issue in laboratory animal science.
Throughout the world, laboratory animal facilities are facing animal welfare issues that come along with housing mice under unnatural conditions. Standard laboratory housing does not reflect social structures of free-living mice, which are comprised of a dominant male, subordinate males, females, and their offspring (Hurst et al. 1993, van Zegeren 1979). A common code of housing practice requires mice to be kept in single-sex groups with few, clearly-defined exceptions, e.g. in the occurrence of deleterious effects due to social incompatibility, veterinary needs, and scientific justification which require single-housing (Kappel et al. 2017, National Research Council (US) 2001). However, group-housing of male mice can cause serious welfare issues. Aggressive territorial and resource-related behaviour among conspecifics leads to stress due to repeated social defeat and subordination and may result in injuries or death (Van Loo et al. 2003, Weber et al. 2017). Since mice are unlikely to adapt to these social stressors, chronic exposure may translate the stress-response into pathological conditions such as immune disorders and increased disease susceptibility (Bartolomucci 2007). When an unacceptable level of aggression is reached (e.g. fights, wounds, body weight loss), male mice are separated and housed individually, which, in turn, diminishes welfare. Deleterious consequences of lack of social contact include increased anxiety- and depression-like behaviours, stress-related corticosterone concentrations, adrenal-body weight-ratios and reduced concentrations of brain-derived neurotropic factor (BDNF) compared to group-housed mice (Berry et al. 2012). The depressive state is accompanied by effects on the serotonergic system (Kalliokoski et al. 2014). Valzelli summarized the changes in behavioural and neurochemical functions of socially deprived male mice under the term “isolation syndrome” (Valzelli 1973).
In the scope of the 3Rs (replace, reduce, refine) by Russel and Burch (Russel and Burch 1959), any contribution to refinement of male mice housing improves welfare (Lloyd et al. 2008). One strategy of eliminating aggression among males is castration. However, the German Animal Welfare Act prohibits the total or partial removal of body parts or the total or partial removal or destruction of organs or tissues of a vertebrate (§ 6 Abs. 1 TierSchG). This prohibition does not apply if the intervention is necessary in individual cases according to veterinary indications and also if sterilisation is carried out in order to prevent uncontrolled reproduction or – unless veterinary considerations prevent it – to continue using or keeping the animal (§ 6 Abs. 1 Nr. 1a, Nr. 5 TierSchG).
The usability of castrated male mice is limited but partial solutions have been implemented by a few laboratory animal facilities, in which castrated male mice are used as sentinels in health-surveillance programs (Lofgren et al. 2012) or for educational purposes (Vaughan et al. 2014) in order to reduce the number of surplus animals.
Only a few studies have examined the benefits of castration in terms of refinement. Castration of CD1 mice at the age of three to four weeks reduced the prevalence of fighting in comparison to intact males from 64 to 0% (Lofgren et al. 2012). Similarly, male outbred Swiss ARC;ARC(S) mice castrated at the age of six to nine weeks showed less agonistic and defensive behaviour compared to intact males and bite injuries were reduced to zero (Vaughan et al. 2014). Common denominators of these studies, however, were the time-point of castration – which was performed at an early age when group housing is still possible (i.e. the level of aggression of the intact males had not required their separation yet) – and the absence of prolonged separation (i.e. mice were group-housed before and after castration). This raises the question whether male mice of an advanced age of more than nine weeks and which had been separated for a prolonged period of time can be resocialized in stable groups after castration. This issue may be especially relevant in scenarios in which a study has been completed and the animal’s general state of health and well-being have been fully restored. If mice are reused (e.g. for educational purposes) in additional research studies or kept as sentinels for a prolonged period of time, castration and resocialization may improve their well-being compared to long-term individual housing.

Themenheft Tiermedizinische Ethik
Description of the case
Top Job:
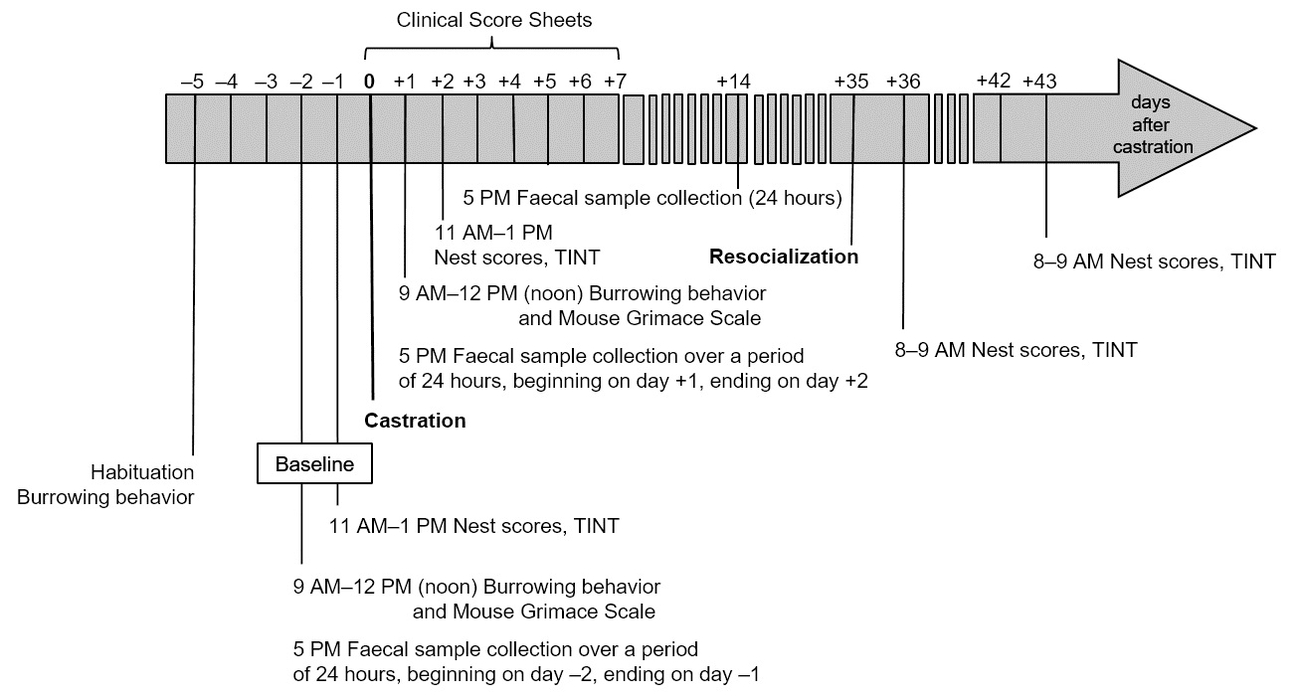
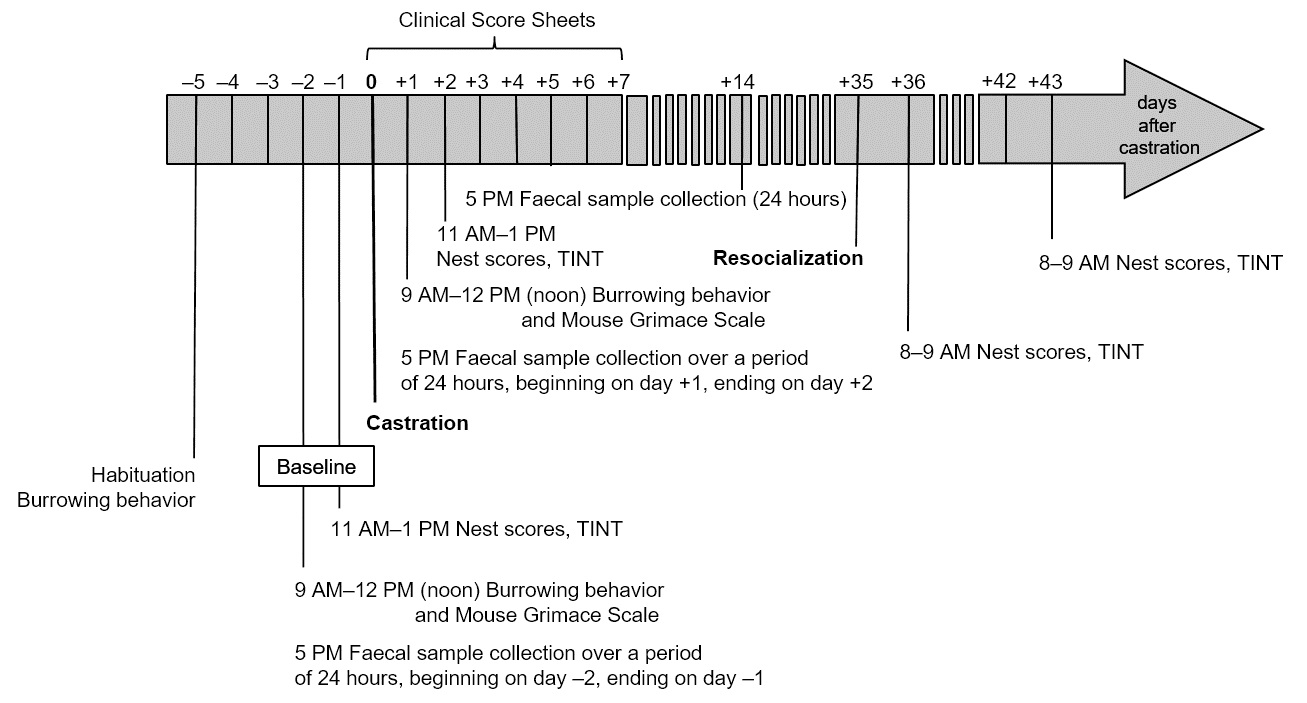
19 male C57BL/6JRj mice from Janvier Labs (Saint-Berthevin Cedex, France) aged 18 to 42 weeks had been used in an experiment which did not necessitate the killing of the animals and their general state of health and well-being had been fully restored. Therefore, humane killing of the mice was not justified. However, the mice had been individually housed for 6 to 32 weeks due to aggressive behaviour. Since long-term individual housing is accompanied by negative effects, the veterinary indication of castration with subsequent resocialization in groups was given. A week post-castration and 28 days prior to resocialization, mice were habituated to the odour of their future group members by distributing nest material among all group members every three to four days. The newly formed groups were comprised of three to four animals. To investigate the impact of surgery on their well-being and whether castration allows for resocialization in stable groups, we analysed two separate time-frames: post-castration and post-resocialization (Fig. 1). First, the well-being of the individual mice post-castration was objectively evaluated by employing behavioural parameters, such as the Mouse Grimace Scale, nest building and burrowing behaviour, biochemical parameters, i.e. concentrations of faecal corticosterone metabolites (FCMs), and changes in body weight. Analgesia was provided by meloxicam (1 mg/kg body weight, s.c. after anaesthesia induction) (Henke et al. 2015) and lidocaine/prilocaine cream (topically applied onto the scrotum). Secondly, the process of resocialization after rehousing in groups of three to four animals at day 35 or day 36 post-castration was carefully analysed. Besides group interaction, group nests and the course of body weight were evaluated in order to assess group stability. Our analyses provided strong indicators of well-being assessment and allowed for a retrospective harm-benefit analysis of castration and resocialization for the 19 mice of this study. A detailed description of materials and methods can be found in the supplementary material.
The authors decided to omit a control group, i.e. resocialization of intact male mice, due to animal welfare reasons and the lack of ethical justification of their expected suffering.
Assessment of well-being after castration
Mouse Grimace Scale (MGS)
The five facial action units (i.e. orbital tightening, nose bulge, cheek bulge, ear position, and whisker change) described in the MGS (Langford et al. 2010) were scored on a scale from 0 to 2 (0 = not present, 1 = moderately present, 2 = obviously present) and, for each scorer, the mean of the five facial action units was calculated. Then mean MGS scores of the two observers were averaged for each mouse for further statistical analysis (Hohlbaum et al. 2017, 2018)
A Pearson correlation was run to determine the relationship between the MGS scores obtained by the two scorers. According to Landis and Koch (1977), there was an almost perfect correlation, which was statistically significant (r = 0.773, p 0.001, n = 133, data of 19 mice obtained at seven time points were included in the analysis).
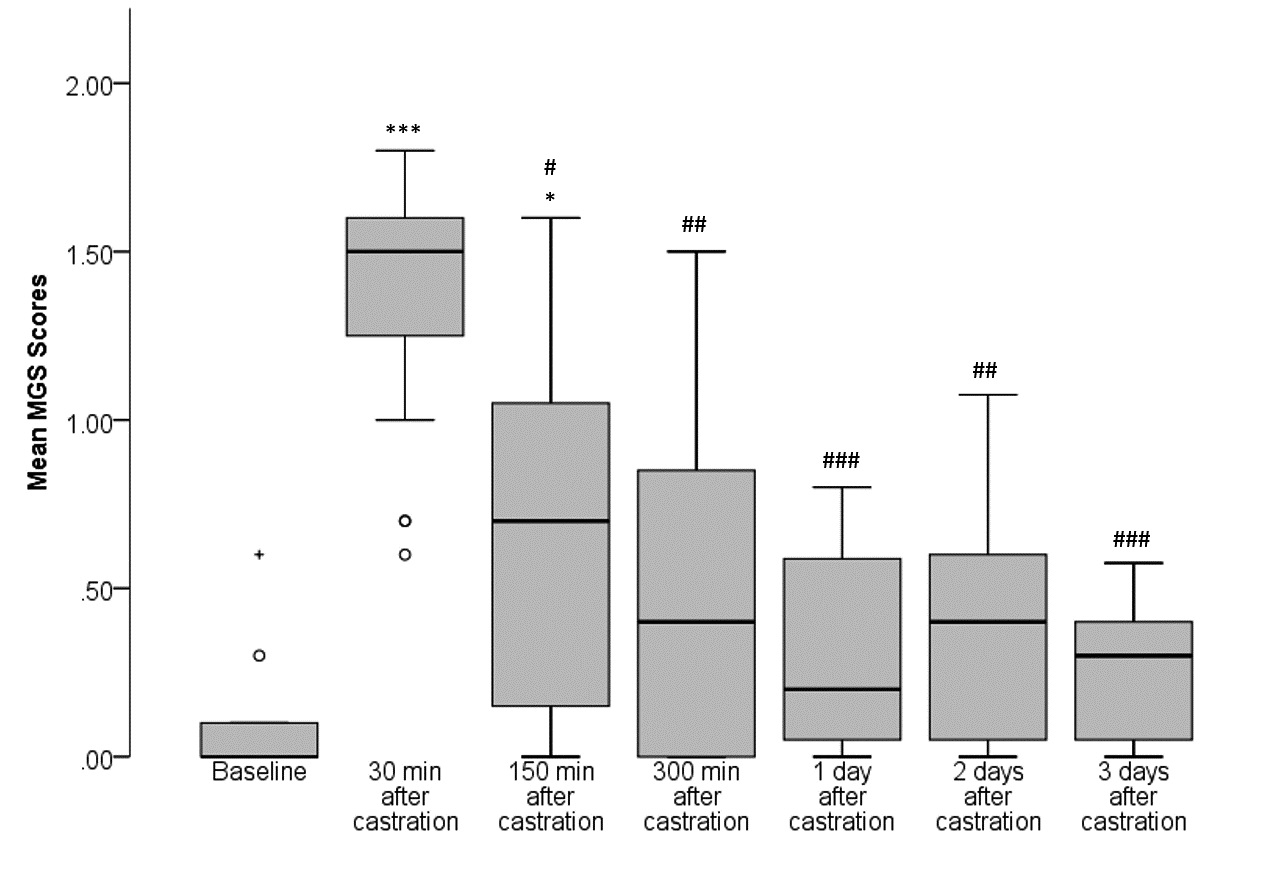
Friedman’s two-way analysis of variance by ranks indicated significant differences between mean MGS scores obtained at the different time points (Chi2 = 53.206, df = 6, p 0.001, n = 19; Fig. 2). The post-hoc Dunn-Bonferroni test revealed increased mean MGS scores at 30 min (p 0.001) and 150 min (p = 0.017) after surgery compared to baseline (i.e. before surgery). Furthermore, mean MGS scores were significantly elevated at 30 min after surgery in comparison to 150 min (p = 0.030), 300 min (p = 0.002), day 1 (p 0.001), day 2 (p = 0.001), and day 3 (p 0.001) after surgery. Data of this figure were reused for another project titled “Towards a fully automated surveillance of well-being status in laboratory mice using deep learning”, which has not been published yet.
Burrowing behaviour
For burrowing behaviour analysis (Deacon 2006, Deacon et al. 2001, Jirkof et al. 2010), a standard opaque plastic water bottle was filled with food pellets and placed in the home cage of the animals. After 2 h, the weight of food pellets removed from the burrow relative to the initial weight was calculated (%).
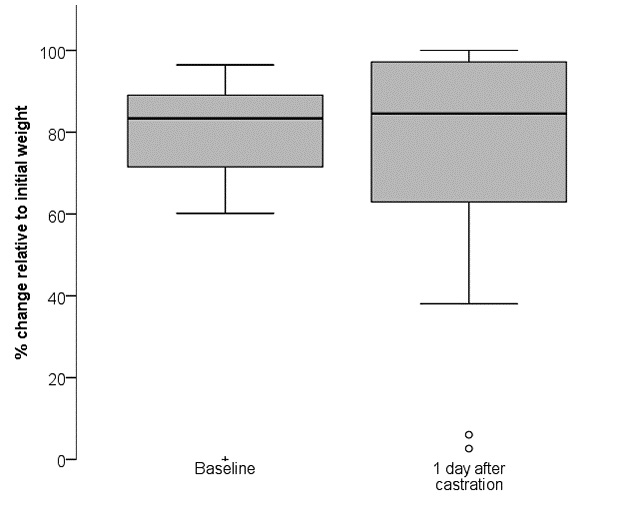
Overall, on day one after castration, burrowing behaviour was not reduced compared to baseline (related samples Wilcoxon signed rank test: z = −0.443, p = 0.658; Fig. 3). At baseline, one mouse did not show burrowing behaviour (0 %) but removed approximately 89 % of the food pellets from the burrow at day one after castration. Two mice showed poorer burrowing performance on day one than at baseline (3 % versus 88 %, 6 % versus 90 %).
Nest building and time-to-integrate-to-nest test (TINT)
Nests were scored on a scale from 0 to 5. In brief, if no nest site was to be identified, the nest score was 0 (undisturbed: mice did not move the material, interact with or manipulate it) or 1 (disturbed: mice interacted with the material but did not use it for nest building). If a nest site was present, the nest was considered to be composed of four parts which were scored individually (i.e. 2 = flat nest, 3 = cup, 4 = incomplete dome, 5 = dome) (Hess et al. 2008).
A Pearson correlation was run to determine the relationship between the nest scores obtained by the two scorers. According to Landis and Koch (1977), there was an almost perfect correlation, which was statistically significant (r = 0.817, p 0.001, n = 38, data of 19 mice obtained at two time points were included in the analysis).
Nest scores obtained at baseline and two days after castration did not significantly differ from each other (related samples Wilcoxon signed rank test: z = 0.240, p = 0.810, Table 1).
A cocoon nestlet piece was placed on the opposite side of the cage from the main nest site and the mice were monitored for 10 min (Rock et al. 2014). The latency to first interaction with novel material as well as the latency to integrate novel material into the present nest were manually recorded.
Wilcoxon signed rank test neither revealed significant differences in the latency to first interaction with the novel material (z = 1.489, p = 0.136), nor in the latency to integrate the novel material to the nest (z = 1.161, p = 0.245) between baseline and day two after castration (Table 1).
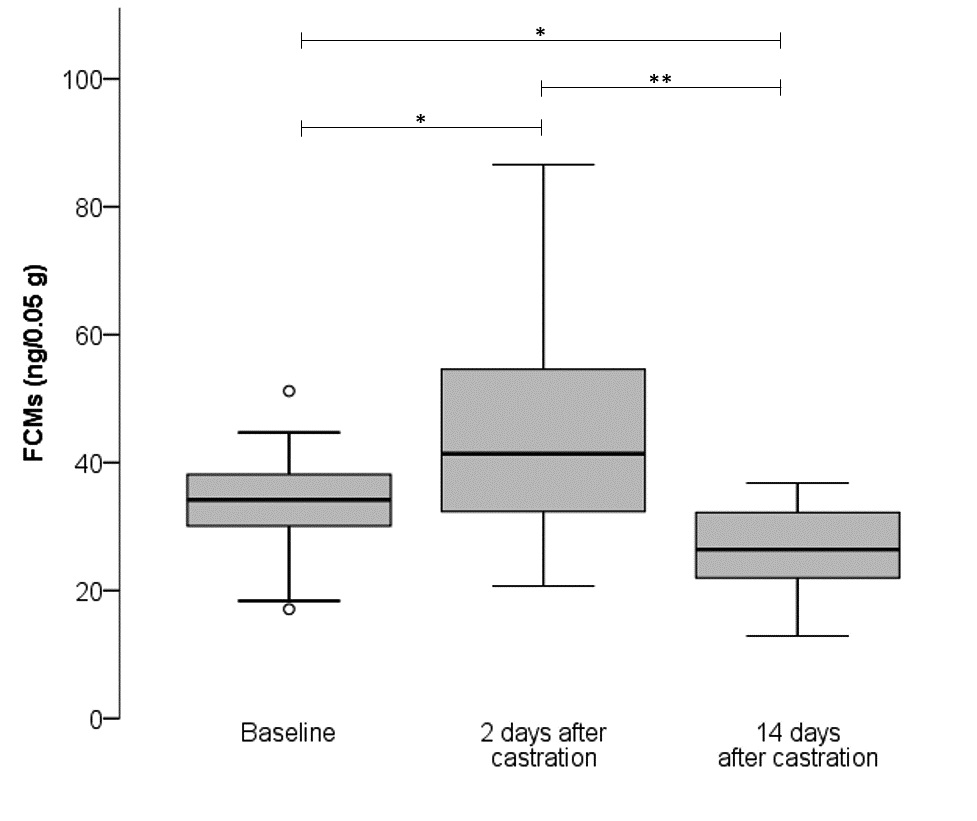
Faecal corticosterone metabolites (FCMs)
Using an ANOVA with repeated measures and a Greenhouse-Geisser correction, the mean values of faecal corticosterone metabolites (FCMs) differed significantly [F (1.324, 23.824) = 15.135, p 0.001; Fig. 4]. Pairwise comparison using Bonferroni post-hoc test revealed significantly elevated FCM concentrations on day two after castration compared to baseline (p = 0.010). Furthermore, FCM concentrations on day 14 after castration were significantly lower than at baseline (p = 0.011) and on day two (p = 0.001).
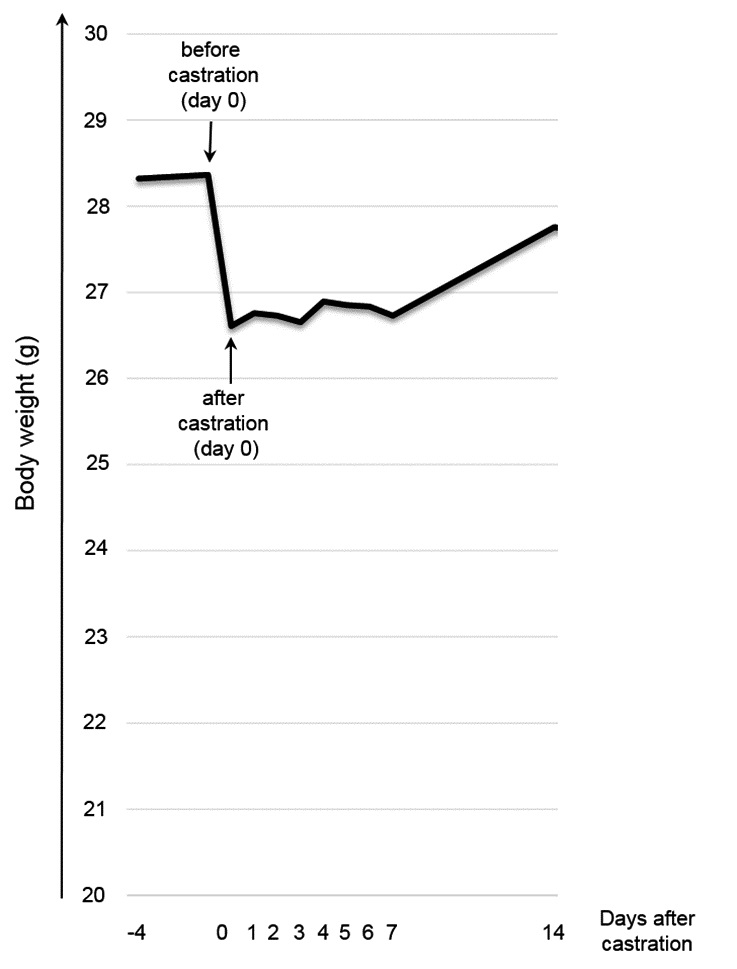
Body weight
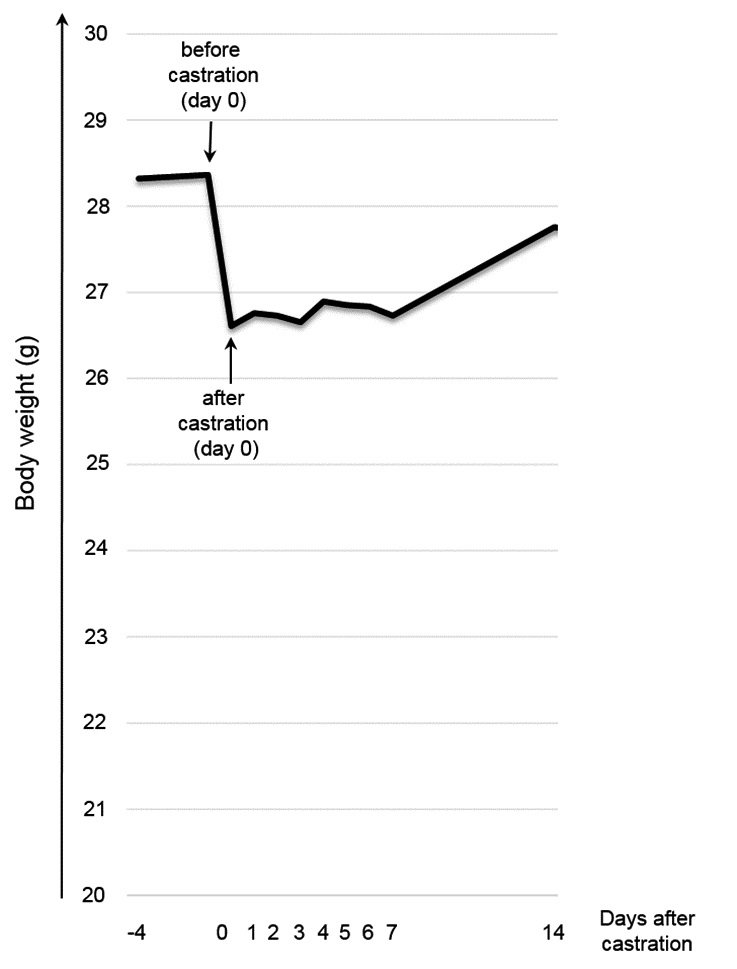
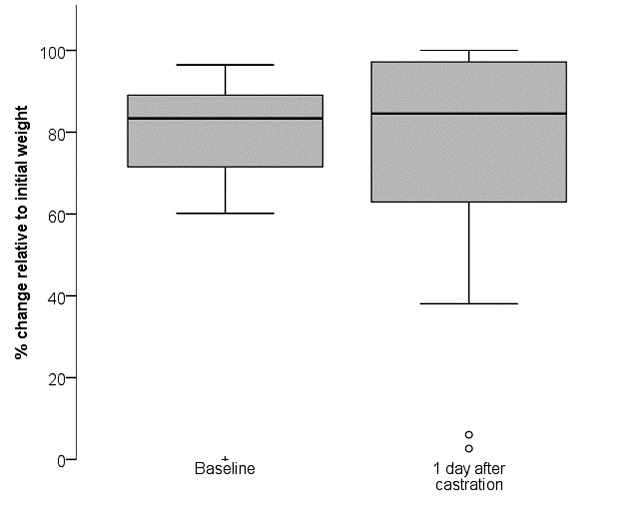
Mice had lost approximately 1.76 g of total body weight on average after surgery on day zero) (Fig. 5), of which 0.48 ± 0.11 g (mean ± standard deviation) are attributed to the tissue removed during surgery. Hence, 1.28 ± 0.95 g of body weight were lost due to post-surgical and/or post-anaesthetic pain and/or distress. Between day zero and day 14, ANOVA with repeated measure and a Greenhouse-Geisser correction revealed a significant gain in body weight [F(3.745, 67.409) = 6.613, p 0.001]. Pairwise comparison by Bonferroni post-hoc indicated significantly higher body weight on day 14 compared to day one to eight after castration (day 1: p = 0.037; day 2: p = 0.001, day 3: p = 0.006, day 4: p = 0.010, day 5: p =0.001, day 6: p = 0.001, day 7: p 0.001, day 8: p 0.001).
Clinical score sheet
A clinical score sheet (Supplementary Table 1) was applied prior to surgery, at 30 min, 150 min, and 300 min after surgery as well as on days one through seven after surgery. It included body posture, coat condition, respiration (pattern), as well as the presence of audible vocalization, pain behaviour, eye closure, diarrhoea, and the condition of the castration wound.
Thirty minutes after surgery, 14 out of 19 mice showed an unsteady, careful gait and reduced rearing, five mice showed reduced forward movement and a hunched posture, and seven mice staggered when moving forward. Furthermore, the coat of 18 mice was slightly unkempt. Two hours later, three mice showed an unsteady, careful gait, as well as reduced rearing and two mice showed a reduced forward movement, whilst two mice still showed staggering and three had a slightly unkempt coat. 300 min after surgery, staggering was no longer observed, however, five mice still showed an impaired body posture (one mouse: slightly unsteady, careful gait, as well as slightly reduced rearing; two mice: unsteady, careful gait, as well as reduced rearing; one mouse: slightly reduced forward movement, slightly hunched back and sunken flanks; one mouse: reduced forward movement, hunched back and sunken flanks) and one mouse had a slightly unkempt coat. On day one after surgery, two mice showed an unsteady, careful gait, as well as reduced rearing and one mouse was dull and slightly unkempt. Audible vocalization, diarrhoea or impaired respiration were not observed at any time point.
Two weeks after surgery, two out of 19 mice developed an inguinal abscess. The abscess was incised, drained, and rinsed with iodine solution under isoflurane anaesthesia. One mouse underwent this procedure only once and was injected subcutaneously (s.c.) with enrofloxacin (10 mg/kg) and meloxicam (1 mg/kg) for five days. For the other mouse, this procedure had to be repeated on four consecutive days. It was injected subcutaneously with enrofloxacin and meloxicam for seven days.
Assessment of well-being post-resocialization
Behavioural analysis of group interaction and lesions resulting from fights
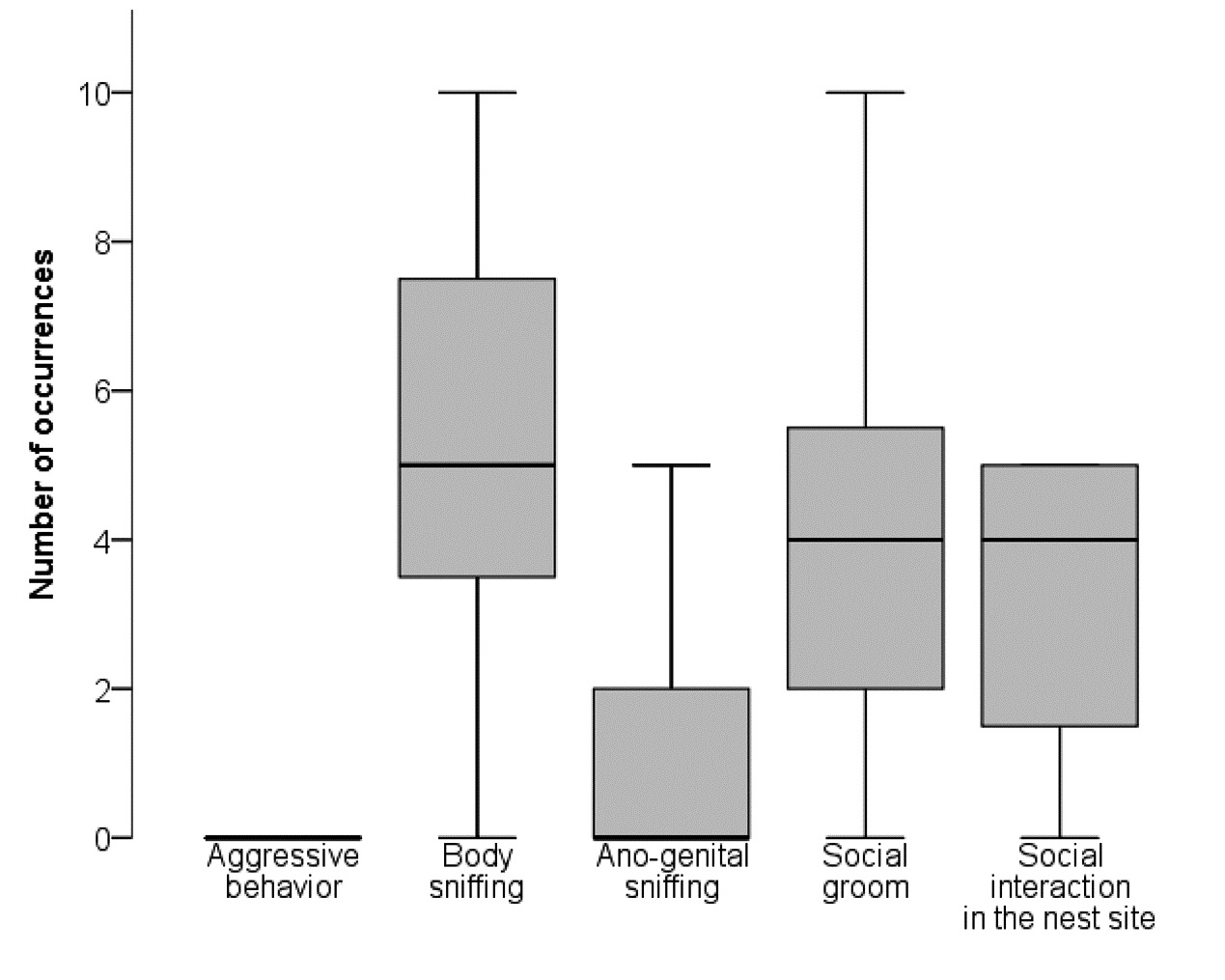
Group interaction was video-recorded for 3 hours. The number of occurrences of behavioural parameters was manually counted during minutes 1–10, 60–65, as well as 120–125 by analysing the videos for each mouse.
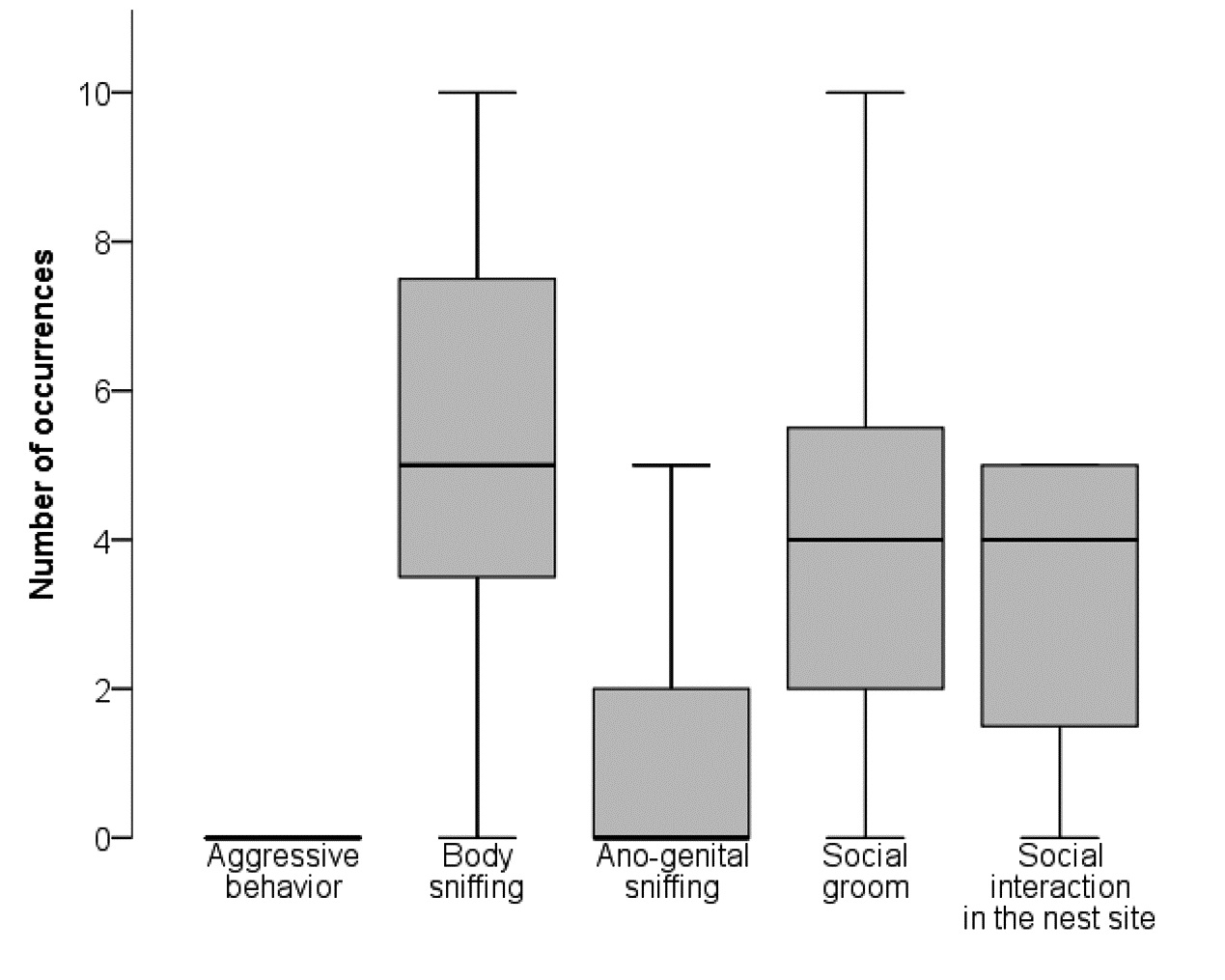
No aggressive behaviour (i.e. chasing, attacking, fighting, biting) was observed during the observation period (i.e. in minutes 1–10, 60–65, and 120–125 after resocialization) (Fig. 6). 18 mice showed body sniffing, eight mice ano-genital sniffing, 16 mice social groom, and 15 mice social interaction in the nest site. The number of occurrences of each behaviour is shown in Fig. 6. No lesions resulting from fights were found during the daily inspection of the mice.
Nest building and time-to-integrate-to-nest test (TINT)
A Pearson correlation was run to determine the relationship between the nest scores obtained by the two scorers. According to Landis and Koch (1977), there was an almost perfect correlation, which was statistically significant (r = 0.709, p = 0.022, n = 10, data of 5 mouse groups obtained at two time points were included in the analysis).
After resocialization, mice built high and complex nests in which they were found to be resting together (Fig. 7). The group nest scores did not significantly differ between day one and day eight after resocialization (related samples Wilcoxon signed rank test: z = −0.944, p = 0.345) (Table 2). On both days and in all five groups, at least one mouse interacted with the novel material and integrated it into the nest. Statistical analysis showed no differences in the latency to first interaction with novel material (related samples Wilcoxon signed rank test: z = −1.753, p = 0.080) and the latency to integrate novel material to nest (related samples Wilcoxon signed rank test: z = −1.214, p = 0.225) between day one and day eight (Table 2).
Body weight after resocialization
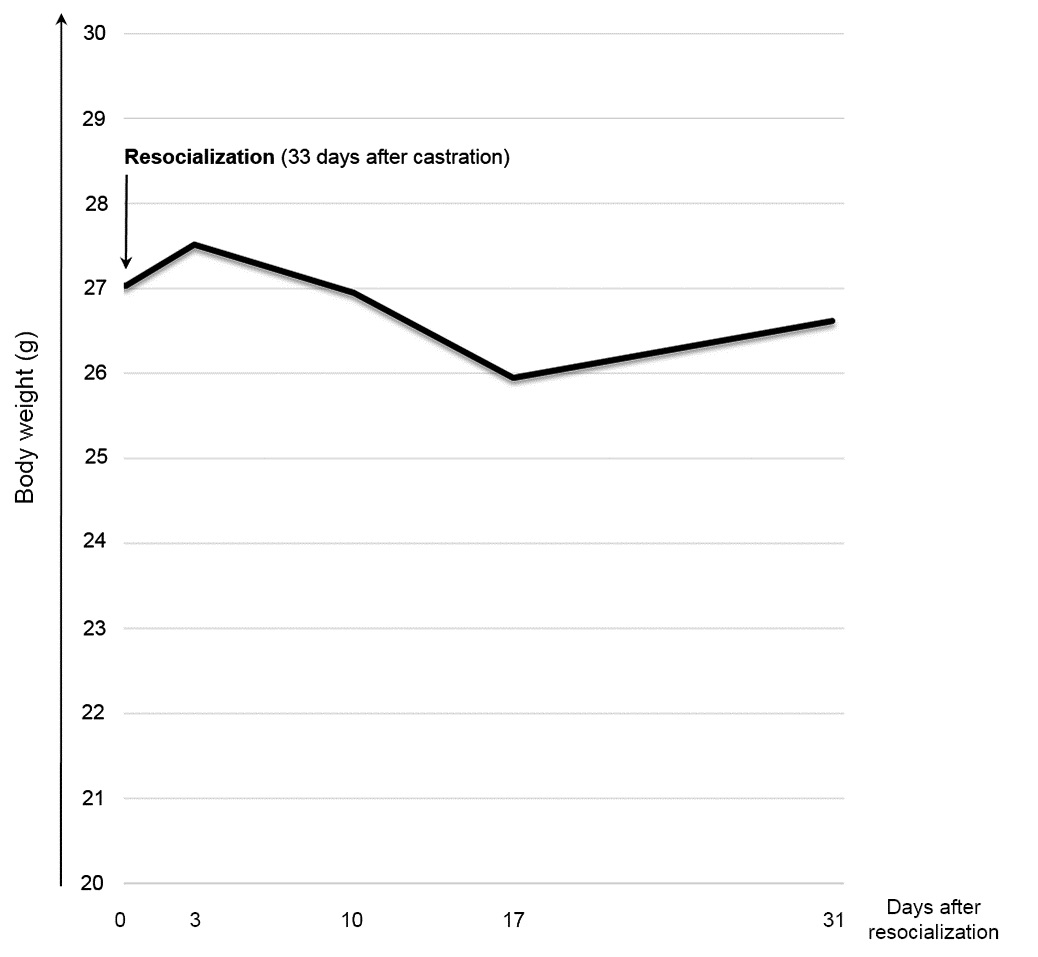
Using ANOVA with repeated measures, a significant difference in body weight over time [F(4, 72) = 21.999, p 0.001, Fig. 8] was observed. Pairwise comparison using Bonferroni post-hoc test revealed an increase in body weight at day three (day 0 versus day 3: p = 0.022) and a loss between day three and 17 after resocialization (day 3 versus day 10: p = 0.025; day 3 versus day 17: p 0.001; day 10 versus day 17: p 0.001). Between day 17 and 31 (end of the study), body weight increased again (p = 0.009).
Discussion
Our observations revealed that resocialization of adult male C57BL/6JRj mice, which had been individually housed for a prolonged period of time of up to 32 weeks, is possible after castration. We closely monitored the phases 1) post-castration (day zero to day 14 post-surgery) and 2) post-resocialization (day 35 to day 56 post-surgery) and used this scientifically sound information as a basis of the harm-benefit analysis, which weighed the benefits of housing castrated male mice in groups against the harm of castration and stressful process of resocialization.
Post-castration
The efficacy of pain management was assessed by the MGS (Langford et al. 2010), a clinical score sheet, burrowing performance, and nesting behaviour. Furthermore, body weight and FCM concentrations were measured as indicators of post-surgical stress.
The MGS scores following castration need to be interpreted with caution since the inhalation anaesthetic isoflurane itself is capable of elevating MGS scores at 30 min post-anaesthesia, whereas its effects are negligible at 150 min (Hohlbaum et al. 2017, Miller et al. 2015). Therefore, the effects of both isoflurane and post-surgical pain may have accumulated at 30 min post-surgery (i.e. 30 min after mice woke up from anaesthesia and showed first forward movements) and resulted in increased MGS scores which, in turn, decreased at 150 min post-surgery but were still elevated compared to baseline. We assumed that the MGS scores reflected the sole impact of post-surgical pain, i.e. excluding effects of isoflurane at this later time point. At 300 min post-surgery, MGS scores were no longer significantly elevated. Overall, however, the MGS scores indicated an ineffectiveness of pain management with standard systemic 1 mg/kg of meloxicam (Henke et al. 2015) and topical lidocaine/prilocaine in male C57BL/6JRj mice undergoing castration via scrotal approach. Furthermore, our observations support the assertion by Miller et al. that “the most critical time period for the provision of analgesia to mice is in the first few hours immediately post-surgery” (Miller et al. 2016). Therefore, the animal should be closely monitored during this period.
Nest building and burrowing behaviour can be impaired by pain or distress (Arras et al. 2007, Jirkof et al. 2010). The high burrowing performance on day one and the construction of complex nests on day two after castration suggest the mice may had already recovered from surgery except for two out of 19 mice. These two mice removed less pellets from the burrow than at baseline, one of which also displayed an increased clinical body posture score (i.e. unsteady, careful gait, as well as reduced rearing), pointing towards the presence of post-surgical pain. Therefore, both mice were additionally treated with one dose of 1 mg/kg meloxicam s.c. on day one. Their burrowing performance was not impaired the following day and the clinical score of the one individual was no longer elevated. It is important to note that these mice are the ones that developed an inguinal abscess two weeks after surgery.
Although the increase in both, the latency to interact with novel material and to integrate it to the nest (TINT), was not statistically significant on day two after castration, it is notable that both parameters were elevated by approximately 1.5 fold when compared to baseline. Whether the parameters obtained by the TINT reflect pain itself or rather non-pain-associated distress remains debatable since the TINT may be affected by diverse negative experiences (Rock et al. 2014). Furthermore, previous studies reported that single housing can lead to inconsistencies in these parameters (Häger et al. 2015, Rock et al. 2014).
FCM concentrations are effective stress markers (Palme 2019) and were assessed by a group-specific enzyme immunoassay validated for mice (Touma et al. 2003, 2004). Since FCM excretion is influenced by the circadian rhythm (Touma et al. 2003, 2004), faecal samples were collected over a period of 24 hours. In the present study, FCM concentrations revealed an elevated HPA-axis activity pointing to post-surgical stress on day two after castration compared to baseline. Using a commercial enzyme immunoassay kit, Wright-Williams et al. (2007) also found elevated FCM concentrations post-vasectomy in male C57BL/6JCrl mice treated either with saline, 5 or 10 mg/kg of meloxicam. Only at a higher dose of 20 mg/kg FCM concentrations were similar to those in untreated animals (Wright-Williams et al. 2007).
Considering the presence of pain-related behaviour assessed by the clinical score sheets, increased MGS scores, and elevated FCM levels during the post-surgical period in our study, pain management by 1 mg/kg meloxicam (Henke et al. 2015) and EMLA cream administered after anaesthesia induction appears not to be sufficiently effective in male C57BL/6JRj mice. It may be improved by a subcutaneous injection of lidocaine/bupivacaine at the incision site before the first cut is made. This should replace the use of EMLA cream, of which onset and duration of action is unclear in mice. Moreover, an intratesticular and/or intrafunicular injection of a local anaesthetic may improve analgesia in mice. This approach has been discussed for the castration of piglets (CASTRUM consortium 2006) in which tearing or cutting the spermatic cord is the most painful procedure of castration (Haga and Ranheim 2005, Taylor and Weary 2000, White et al. 1995). However, the risk of systemic effects following the absorption of high concentrations of local anaesthetics from the injection site into the blood stream must also be considered (Becker and Reed 2012) and should be further investigated. If the improved local anaesthesia does not decrease post-surgical pain and distress, the dose of meloxicam may be increased (Wright-Williams et al. 2007) (e.g. 5 mg/kg) and injected every 12 hours. Another option may be the combination of a non-steroidal anti-inflammatory drug (NSAID) with buprenorphine (Wright-Williams et al. 2013). Preemptive analgesia could be provided by voluntary oral administration of buprenorphine or a NSAID, e.g. via (sweetened) drinking water (Jirkof et al. 2019) in the dark phase prior to surgery. To ensure adequate intake of analgesics, the mixture of analgesics and sweetened drinking water can alternatively be offered in a syringe for voluntary consumption (Lang 2016). Moreover, analgesics can be mixed with flavoured jelly (Liles et al. 1998) or nut paste (Abelson et al. 2012). However, voluntary oral administration requires the habituation of the mice to the novel food before the procedure in order to prevent neophobia.
Interestingly, FCM levels were lower at day 14 after castration compared to day two post-surgery and also to baseline which may be caused by the lack of endogenous, testicular testosterone due to the removal of both testes and associated changes in the endocrine system. Faecal testosterone metabolites were already decreased on day 2 after castration (Auer et al. 2020). It had previously been shown that castration increased adrenal 4α-reductase activity which resulted in a reduction of corticosterone and an increase of adrenal corticosterone metabolites (Colby and Kitay 1972). In contrast, Kalil et al. (2013) demonstrated that orchiectomy did not alter basal or stress-induced blood corticosterone concentrations in male Wistar rats on day 11 after castration. However, our FCM analysis showed a decrease in HPA-axis activity 14 days after castration and suggests decreased stress levels in mice following castration. Stress levels of intact male mice can be triggered by several factors such as the presence of androgen-dependent pheromones in the urine of other male mice, increasing aggressive behaviour (Chamero et al. 2007, Lee and Wilson 2012, Mugford and Nowell 1970). The mice in the present study were kept under housing conditions permitting continuous exposure to the odours of intact males, which may have resulted in higher stress levels. Castrated male mice do not produce such pheromones and, therefore, fail to induce aggressive behaviour (Mugford and Nowell 1970). This is in line with the reduced FCM excretion in castrated mice of our study.
The increase in body weight supports our hypothesis that the stress levels began decreasing two weeks after castration, but it remains to be clarified whether the increase may also be explained by the continuous weight gain over time until mice reach advanced age (Fahlström et al. 2011). In C57BL/6NJ mice, it was shown that body weight increased until 15–20 months of age (Fahlström et al. 2011). However, another study reported that castrated C57BL/6J mice gain less weight over time than intact mice since calorie intake (i.e. food intake) declines whereas cumulated feed efficiency (body weight gain/calorie intake) is unaltered (Harada et al. 2016).
All in all, our investigations conducted post-castration indicate that surgical intervention caused short-term pain (i.e. until at least 150 min post-surgery) and stress (until at least 2 days post-surgery), except in the two mice that developed an inguinal abscess and had to be treated accordingly. An improved analgesic regimen may ameliorate the degree of stress and pain the mice experienced during the early post-surgical period.
Post-resocialization
We demonstrate that castrated male C57BL/6JRj mice can be resocialized in same-sex groups of three to four mice. Neither signs of aggressive behaviour during the observation period immediately after resocialization nor fight-associated lesions during the entire observation period of three weeks after resocialization occurred. It should be noted that mice were habituated to the odour of their future group members by distributing nest material among all group members every three to four days, beginning approximately a week post-castration.
On day one as well as on day eight, all groups built high and complex nests indicative of group stability (Hayes 2000, Van Loo et al. 2003) since instability triggers fighting which, in turn, leads to nests of poorer quality and/or destruction of nests. Further, novel nest material was integrated into the nest within approximately 2 min on day one and more quickly, i.e. 1 min, on day eight after resocialization, which reflects the effort of the mice to build complex group nests and is mirrored by the higher nest complexity scores.
Body weight decreased within the first two weeks after resocialization and increased within the following two weeks. The course of body weight may reflect the process of habituation to the new group, which initially may have caused stress influencing body weight. For a deeper insight into the actual stress levels, FCM should be analysed at several time points after resocialization in future studies.
Our study is based on two previous investigations in which castration of male CD1 mice three to four weeks of age (Lofgren et al. 2012) and outbred Swiss ARC;ARC(S) mice six to nine weeks of age (Vaughan et al. 2014) reduced aggressive behaviour and improved group-housing. It is important to note that both studies evaluated the effects of castration in group-housed male mice of up to nine weeks of age. Age and (group) housing conditions are the major aspects in which our study design differs from Lofgren et al. (2012) and Vaughan et al. (2014): the C57BL/6JRj mice in our study were 18–42 weeks of age and had been individually housed for a prolonged period of time of up to
32 weeks due to conspecific aggressive behaviour.
Nevertheless, our findings demonstrate that resocialization in stable groups without the occurrence of fight-associated lesions due to aggressive behaviour is possible after castration.
Harm-benefit analysis
Within the framework of the Netzwerk Tiermedizinische Ethik (Network of Veterinary Ethics), which combines ethics and veterinary medicine and aims to establish the research field of veterinary ethics (Weich 2019), the authors discussed the present dilemma: The harm-benefit analysis focuses on the dilemma whether it is favourable to single-house or castrate aggressive male mice, by which latter allows for resocialization and harmonious group housing. The benefits resulting from castration and resocialization only become relevant if the improvement of well-being outweighs the harm incurred by surgery.
Co-housing intact males can result in deleterious consequences due to aggressive territorial and resource-related behaviour, leading to stress or even fatal painful injuries. This diminishes and, hence, does not seem to comply with animal welfare. In order to protect the mice from these deleterious consequences, they are often housed individually, although the lack of social contact can increase distress and anxiety and depression-like behaviours (Berry et al. 2012, Kalliokoski et al. 2014, Valzelli 1973). Taken together, well-being of intact male mice can be significantly impaired by both group- and single-housing.
Castration is accompanied by short-term pain and distress, as demonstrated by our investigations conducted post-castration. The degree of pain and distress found in the present case report may be ameliorated by improved analgesic regimen. If complications occur after surgery (i.e. inguinal abscesses), the degree and duration of pain and distress will increase and impair the welfare of the respective animal. Thorough post-surgical care and observation of species-specific pain behaviour is required to identify these animals and treat them appropriately. However, these treatments may also cause additional stress, which can be minimized by using the route of application that induces as little stress as possible (e.g. voluntary oral intake, sustained release formulations). Castration can also cause long-term impairment due to elimination of male sex hormones (Ayaz et al. 2019) despite allowing for prolonged housing in stable single-sex male groups. Moreover, the habituation phase to the new group can represent a stressor itself.
Regarding legal terms and definitions, the German Animal Welfare Act generally prohibits the removal of organs of vertebrates. However, a legal exemption is provided for procedures which are based on veterinary indication and is not limited to curative treatment of physical pathologies (§ 6 Abs. 1 Nr. 1a TierSchG). Another exemption is sterilisation in order to prevent uncontrolled reproduction or to continue using or keeping the animal (§ 6 Abs. 1 Nr. 5 TierSchG). In the case of male mice showing aggressive behaviour, which is likely caused by sexual hormones and can lead to physical or mental suffering, castration may be an option, provided that a competent veterinary assessment has been made. The Tierärztliche Vereinigung für Tierschutz e.V. (TVT, Veterinary Association for the Protection of Animals) published a guideline on the castration of cats and dogs which states castration as appropriate measure for shelters in maintaining male dogs in harmonious groups (TVT 2011). Extrapolation of this guideline to laboratory animal facilities which are explicitly requested to house mice in social groups may therefore even imply a demand for castrating male mice. However, as previously described in detail by Palmer et al. (2012), routine castration is not morally justified.
From a veterinary perspective, considering that the author’s moral perspective cannot be disregarded, castration of male mice may be the most favourable option in resolving the present ethical dilemma, provided that the principles of good veterinary practice are adhered to. In terms of a lifetime harm-benefit analysis for the individual animal, the short-term post-surgical pain and stress may outweigh the long-lasting negative effects of aggression occurring in single-sex male group-housing for a prolonged period of time and also to the consequences of social isolation associated with long-term individual housing. To prevent any surgical complications, that may increase the degree and duration of post-surgical pain and stress, proper pain treatment and thorough post-surgical care must be ensured. Moreover, observation of species-specific pain behaviour up to several weeks after castration is absolutely essential.
The authors do not suggest castration of aggressive male mice as routine treatment but rather focus on the circumstances present in this particular case, which may be extrapolated to cases of similar nature. Castrated and resocialized mice may be (re)used for educational purposes or as sentinels. With some caution, it may also be considered to use them as social partners for intact or vasectomized mice, though the authors have no experiences with these group compositions. Moreover, it may be discussed to use castrated male mice in long-term studies, in which the animals would otherwise have to be kept socially isolated due to aggression. However, the impact of the altered hormone balance on the study results is a highly relevant issue to discuss and must be considered. Another option applying to the mice in the present case report is rehoming. However, rehoming in a husbandry system suitable for the species can only be considered if the health state of the animals allows for it, the animals do not pose a risk to the health of humans or other animals or to the environment, and appropriate measures have been taken to ensure the welfare of the animals (§ 10 Abs. 1 TierSchVersV). Moreover, it has to be ensured that the animals to be rehomed undergo a habituation process (§ 10 Abs. 2 TierSchVersV). These requirements were met for the mice of the present case report, which had been previously used in an experiment classified as “mild” with regards to Directive 2010/63/EU. The experiment did not necessitate the death of the animals for further investigations. Their general state of health had not been impaired by the previous experiment and their well-being had been fully restored when the study was completed. Due to the previous study, including an extensive behavioural test battery, the mice were habituated to the interaction with humans. Since they had been handled by a combination of tunnel and cup method, anxiety-related behaviour in anticipation of handling was assumed to be very low (Hurst and West 2010). A great benefit was the opportunity to cooperate with an animal welfare organization, which supports rehoming of laboratory animals (e.g. contact with potential new animal owner, inspection or assessment of husbandry systems, transport, etc.). As these requirements are often not met and there are probably only few persons willing to keep laboratory mice as pets, rehoming may only be considered in individual cases.
Taken together, the requirements that need to be observed when castrating male mice and the limitations in the use of castrated male mice emphasize that the harm-benefit analysis must be carried out on a case-by-case basis.
Conclusion
Adult male C57BL/6JRj mice, which had been individually housed for a prolonged period of time due to aggressive behaviour towards their conspecifics, can be resocialized and form stable groups of three to four animals after castration. The surgical intervention causes short-term stress and pain, which may be ameliorated by an improved analgesic regimen.
The surgical intervention creates ethical and animal welfare issues which need to be addressed in veterinary ethics and considered on a case-by-case basis. Taking into account the lifetime experience of the individual animal in this case report, the short-term post-surgical stress and pain may be preferential to the longer lasting negative effects caused by single-housing. Although usability of castrated mice is limited, hygiene monitoring or educational purposes may pose options to reduce the number of surplus animals being humanely killed. In individual cases, it may be considered to rehome or use castrated male mice in long-term studies.
Acknowledgments
We thank Sabine Jacobs for assisting with the faecal sample collection and MGS scoring, Dr. Silke Dietze for scoring the nests, Edith Klobetz-Rassam for FCM analysis, and PD Dr. med. vet. habil. Roswitha Merle for assisting with statistical analysis.
Conflict of interest
There are no protected, financial, professional or other personal interests in a product, service and/or company that could influence the content or opinions expressed in the above manuscript.
Ethical approval
The study was performed according to the German Animal Welfare Act and the Directive 2010/63/EU for the protection of animals used for scientific purposes. Animal housing and husbandry were approved by the Berlin State Authority (Landesamt für Gesundheit und Soziales, permit number: G 0053/15). Mice had been previously used in an experiment classified as “mild” with regards to animal suffering as defined by current EU regulations on severity classification of animal studies. After castration and resocialization in groups of three to four, the mice were rehomed.
Funding
This work was not supported by a grant.
Authors’ contribution
Conception or design of the work: KH, CL (KH and CL contributed equally to this work); data collection: KH; data analysis and interpretation: KH; drafting the article: KH; critical revision of the article: KH, CL, RP, NAE, NK, NB, JB, CTR (JB and CTR contributed equally to this work); final approval of the version to be published: KH, CL, RP, NAE, NK, NB, JB, CTR.
Electronic supplementary material
A detailed description of materials and methods can be found in the supplementary material document.
Addresses for correspondence
Katharina Hohlbaum
Freie Universität Berlin
Department of Veterinary Medicine
Institute of Animal Welfare, Animal Behavior, and Laboratory Animal Science
Königsweg 67 (Haus 21)
14163 Berlin, Germany
Katharina.hohlbaum@fu-berlin.de
Charlotte Seibert (geb. Leidinger)
Translational Animal Research Center (TARC)
Universitätsmedizin der Johannes Gutenberg-Universität Mainz
Hanns-Dieter-Hüsch-Weg 19
55128 Mainz, Germany
Charlottesophieseibert@gmail.com
References
Abelson KS, Jacobsen KR, Sundbom R, Kalliokoski O, Hau J (2012): Voluntary ingestion of nut paste for administration of buprenorphine in rats and mice. Lab Anim 46(4): 349–351.
Andresen N, Wöllhaf M, Hohlbaum K, Lewejohann L, Hellwich O, Thöne-Reineke C, Belik V (2020): Towards a fully automated surveillance of well-being status in laboratory mice using deep learning: starting with facial expression analysis. PLoS ONE 15(4): e0228059. https://doi.org/10.1371/journal.pone.0228059.
Arras M, Rettich A, Cinelli P, Kasermann HP, Burki K (2007): Assessment of post-laparotomy pain in laboratory mice by telemetric recording of heart rate and heart rate variability. BMC Vet Res 3: 16.
Auer KE, Kußmaul M, Möstl E, Hohlbaum K, Rülicke T, Palme R (2020): Measurement of fecal testosterone metabolites in mice: replacement of invasive techniques. Animals 10(1): 165.
Ayaz O, Banga SE, Heinze-Milne S, Rose RA, Pyle WG, Howlett SE (2019): Long term testosterone deficiency modifies myofilament and calcium handling proteins and promotes diastolic dysfunction in the aging mouse heart. Am J Physiol Heart Circ Physiol 316: H768–H780.
Bartolomucci A (2007): Social stress, immune functions and disease in rodents. Front Neuroendocrinol 28(1): 28–49.
Becker DE, Reed KL (2012): Local anesthetics: review of pharmacological considerations. Anesth Prog 59(2): 90–102.
Berry A, Bellisario V, Capoccia S, Tirassa P, Calza A, Alleva E, Cirulli F (2012): Social deprivation stress is a triggering factor for the emergence of anxiety- and depression-like behaviours and leads to reduced brain BDNF levels in C57BL/6J mice. Psychoneuroendocrinology 37(6): 762–772.
CASTRUM consortium (2006): Pig castration: methods of anaesthesia and analgesia for all pigs and other alternatives for pigs used in traditional products. Available from: https://op.europa.eu/en/publication-detail/-/publication/5fe8db00-dbb8-… (last accessed on January 8, 2020). DOI 10.2875/057159.
Chamero P, Marton TF, Logan DW, Flanagan K, Cruz JR, Saghatelian A, Cravatt BF, Stowers L (2007): Identification of protein pheromones that promote aggressive behaviour. Nature 450(7171): 899–902.
Colby HD, Kitay JI (1972): Effects of gonadal hormones on adrenocortical secretion of 5 -reduced metabolites of corticosterone in the rat. Endocrinology 91(6): 1523–1527.
Directive 2010/63/EU (2010): Directive of the European parliament and of the council of 22 September 2010 on the protection of animals used for scientific purposes. Official Journal of the European Union. ABI. L276/33-L276/79. https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2010:276:00… (last accessed on January 8, 2020).
Deacon RM (2006): Burrowing in rodents: a sensitive method for detecting behavioral dysfunction. Nat Protoc 1(1): 118–121.
Deacon RM, Raley JM, Perry VH, Rawlins JN (2001): Burrowing into prion disease. Neuroreport 12(9): 2053–2057.
Fahlström A, Yu Q, Ulfhake B (2011): Behavioral changes in aging female C57BL/6 mice. Neurobiol Aging 32(10): 1868–1880.
Grimm H, Bremhorst A, Ach JS (2018): Tierversuche. In: Ach JS, Borchers D (Hrsg.), Handbuch Tierethik. Grundlagen – Kontexte – Perspektiven, Springer-Verlag, Stuttgart, 273–278.
Grimm H (2014): Ethics in Laboratory Animal Science. In: Jensen-Jarolim E (Hrsg.): Comparative medicine: anatomy and physiology, Springer-Verlag, Wien, 281–300.
Grimm H, Olsson IAS, Sandøe P (2019): Harm-benefit analysis – what is the added value? A review of alternative strategies for weighing harms and benefits as part of the assessment of animal research. Lab Anim 53(1):17–27.
Haga HA, Ranheim B (2005): Castration of piglets: the analgesic effects of intratesticular and intrafunicular lidocaine injection. Vet Anaesth Analg 32(1): 1–9. Häger C, Keubler LM, Biernot S, Dietrich J, Buchheister S, Buettner M, Bleich A (2015): Time to integrate to nest test evaluation in a mouse DSS-colitis model. PLoS One 10(12): e0143824.
Harada N, Hanaoka R, Horiuchi H, Kitakaze T, Mitani T, Inui H, Yamaji R (2016): Castration influences intestinal microflora and induces abdominal obesity in high-fat diet-fed mice. Sci Rep 6: 23001.
Hayes LD (2000): To nest communally or not to nest communally: a review of rodent communal nesting and nursing. Anim Behav 59(4): 677–688.
Henke J, Haberstroh J, Sager M, Becker K, Eberspächer E, Bergadano A, Zahner D, Arras M (2015): Fachinformation aus dem Ausschuss für Anästhesie der GV-SOLAS in Zusammenarbeit mit dem Arbeitskreis 4 in der TVT: Schmerztherapie bei Versuchstieren. Available from: www.gv-solas.de/fileadmin/user_upload/pdf_publikation/Anaest._Analgesie… (last accessed on January 8, 2020).
Hess SE, Rohr S, Dufour BD, Gaskill BD, Pajor EA, Garner JP (2008): Home improvement: C57BL/6J mice given more naturalistic nesting materials build better nests. J Am Assoc Lab Anim Sci 47(6): 25–31.
Hohlbaum K, Bert B, Dietze S, Palme R, Fink H, Thöne-Reineke C (2017): Severity classification of repeated isoflurane anesthesia in C57BL/6JRj mice-Assessing the degree of distress. PLoS One 12(6): e0179588.
Hohlbaum K, Bert B, Dietze S, Palme R, Fink H, Thöne-Reineke C (2018): Systematic assessment of well-being in mice for procedures using general anesthesia. J Vis Exp 133: e57046.
Hurst JL, Fang J, Barnard CJ (1993): The role of substrate odours in maintaining social tolerance between male house mice, Mus musculus domesticus. Animal Behaviour 45(5): 997–1006.
Hurst JL, West RS (2010): Taming anxiety in laboratory mice. Nat Methods 7(10): 825–826.
Jirkof P, Cesarovic N, Rettich A, Nicholls F, Seifert B, Arras M (2010): Burrowing behavior as an indicator of post-laparotomy pain in mice. Front Behav Neurosci 4: 165.
Jirkof P, Durst M, Klopfleisch R, Palme R, Thöne-Reineke C, Buttgereit F, Schmidt-Bleek K, Lang A (2019): Administration of tramadol or buprenorphine via the drinking water for post-operative analgesia in a mouse-osteotomy model. Sci Rep 9(1): 10749.
Kalil B, Leite CM, Carvalho-Lima M, Anselmo-Franci JA (2013): Role of sex steroids in progesterone and corticosterone response to acute restraint stress in rats: sex differences. Stress 16(4): 452–460.
Kalliokoski O, Teilmann AC, Jacobsen KR, Abelson KSP, Hau J (2014): The lonely mouse – single housing affects serotonergic signaling integrity measured by 8-OH-DPAT-induced hypothermia in male mice. PLoS One 9(12): e111065.
Kappel S, Hawkins P, Mendl MT (2017): To group or not to group? Good practice for housing male laboratory mice. Animals (Basel) 7(12): 88.
Landis JR, Koch GG (1977): The measurement of observer agreement for categorical data. Biometrics 33: 159–174.
Lang A, Schulz, Ellinghaus A, Schmidt-Bleek K (2016): Osteotomy models–the current status on pain scoring and management in small rodents. Lab Anim 50(6): 433–441.
Langford DJ, Bailey AL, Chanda ML, Clarke SE, Drummond TE, Echols S, Glick S, Ingrao J, Klassen-Ross T, Lacroix-Fralish ML, Matsumiya L, Sorge RE, Sotocinal SG, Tabaka JM, Wong D, van den Maagdenberg AM, Ferrari MD, Craig KD, Mogil JS (2010): Coding of facial expressions of pain in the laboratory mouse. Nat Methods 7(6): 447–449.
Lee DL, Wilson JL (2012): Urine from sexually mature intact male mice contributes to increased cardiovascular responses during free-roaming and restrained conditions. ISRN Vet Sci 2012: 185461.
Liles JH, Flecknell PA, Roughan J, Cruz-Madorran I (1998): Influence of oral buprenorphine, oral naltrexone or morphine on the effects of laparotomy in the rat. Lab Anim 32(2): 149–161.
Lloyd MH, Foden BW, Wolfensohn SE (2008): Refinement: promoting the three Rs in practice. Lab Anim 42(3): 284–293.
Lofgren JL, Erdman SE, Hewes C, Wong C, King R, Chavarria TE, Discua AR, Fox JG, Maurer KJ (2012): Castration eliminates conspecific aggression in group-housed CD1 male surveillance mice (Mus musculus). J Am Assoc Lab Anim Sci 51(5): 594–599.
Miller A, Kitson G, Skalkoyannis B, Leach M (2015): The effect of isoflurane anaesthesia and buprenorphine on the mouse grimace scale and behaviour in CBA and DBA/2 mice. Appl Anim Behav Sci 172: 58–62.
Miller AL, Kitson GL, Skalkoyannis B, Flecknell PA, Leach MC (2016): Using the mouse grimace scale and behaviour to assess pain in CBA mice following vasectomy. Appl Anim Behav Sci 181: 160–165.
Mugford RA, Nowell NW (1970): Pheromones and their effect on aggression in mice. Nature 226(5249): 967–968.
National Research Council, US (2001): Guide for the Care and Use of Laboratory Animals. 8th ed., available from: https://grants.nih.gov/grants/olaw/guide-for-the-care-and-use-of-labora… (last accessed on January 8, 2020).
Palme R (2019): Non-invasive measurement of glucocorticoids: advances and problems. Physiol Beh 199: 229–243.
Palmer C, Corr S, Sandøe PJA (2012): Inconvenient desires: should we routinely neuter companion animals? Anthrozoos 25(sup1): s153–s172.
Rock ML, Karas AZ, Rodriguez KB, Gallo MS, Pritchett-Corning K, Karas RH, Aronovitz M, Gaskill BN (2014): The time-to-integrate-to-nest test as an indicator of wellbeing in laboratory mice. J Am Assoc Lab Anim Sci 53(1): 24–28.
Russel W, Burch R (1959): The principles of humane experimental technique. Methuen, London.
Taylor AA, Weary DM (2000): Vocal responses of piglets to castration: identifying procedural sources of pain. Appl Anim Behav Sci 70(1): 17–26.
The Commission of the European Communities (2007): European Union Commission recommendation of 18 June 2007 on guidelines for the accommodation and care of animals used for experimental and other scientific purposes. Available from: eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32007H0526&from=EN (last accessed on January 8, 2020).
TierSchG (2019): Tierschutzgesetz in der Fassung der Bekanntmachung vom 18. Mai 2006 (BGBl. I S. 1206, 1313), das zuletzt durch Artikel 101 des Gesetzes vom 20. November 2019 (BGBl. I S. 1626) geändert worden ist.
TierSchVersV (2015): Tierschutz-Versuchstierverordnung vom 1. August 2013 (BGBl. I S. 3125, 3126), die zuletzt durch Artikel 394 der Verordnung vom 31. August 2015 (BGBl. I S. 1474) geändert worden ist.
TVT (2011): Tierärztliche Vereinigung für Tierschutz e.V. Merkblatt 120: Kastration von Hunden und Katzen. Available from: www.tierschutz-tvt.de/index.php?id=50&no_cache=1&download=TVT-MB_120_Ka… (last accessed on January 8, 2020).
Touma C, Palme R, Sachser N (2004): Analyzing corticosterone metabolites in fecal samples of mice: a noninvasive technique to monitor stress hormones. Horm Behav 45(1): 10–22.
Touma C, Sachser N, Mostl E, Palme R (2003): Effects of sex and time of day on metabolism and excretion of corticosterone in urine and feces of mice. Gen Comp Endocrinol 130(3): 267–278.
UK Home Office (2014): Code of practice for the housing and care of animals bred, supplied or used for scientific purposes. Rodents and Rabbits. Available from: www.gov.uk/government/uploads/system/uploads/attachment_data/file/38980… (last accessed on January 8, 2020).
Valzelli L (1973): The “isolation syndrome” in mice. Psychopharmacologia 31(4): 305–320.
Van Loo PL, Van Zutphen LF, Baumans V (2003): Male management: coping with aggression problems in male laboratory mice. Lab Anim 37(4): 300–313.
van Zegeren K (1979): Variation in aggressiveness and the regulation of numbers in house mouse populations. Netherlands Journal of Zoology 30(4): 635–770.
Vaughan LM, Dawson JS, Porter PR, Whittaker AL (2014): Castration promotes welfare in group-housed male Swiss outbred mice maintained in educational institutions. J Am Assoc Lab Anim Sci 53(1): 38–43.
Weber EM, Dallaire JA, Gaskill BN, Pritchett-Corning KR, Garner JP (2017): Aggression in group-housed laboratory mice: why can’t we solve the problem? Lab Anim 46: 157–161.
Weich K (2019): Towards veterinary ethics (2010). ALTEX Proceedings 8(1): 218. https://eusaat-congress.eu/images/2019/pdf/altex_Linz2019_full.pdf (last accessed December 20, 2019).
White RG, DeShazer JA, Tressler CJ, Borcher GM, Davey S, Waninge A, Parkhurst AM, Milanuk MJ, Clemens ET (1995): Vocalization and physiological response of pigs during castration with or without a local anesthetic. J Anim Sci 73(2): 381–386.
Wright-Williams S, Flecknell PA, Roughan JV (2013): Comparative effects of vasectomy surgery and buprenorphine treatment on faecal corticosterone concentrations and behaviour assessed by manual and automated analysis methods in C57 and C3H mice. PLoS One 8(9): e75948.
Wright-Williams SL, Courade JP, Richardson CA, Roughan JV, Flecknell PA (2007): Effects of vasectomy surgery and meloxicam treatment on faecal corticosterone levels and behaviour in two strains of laboratory mouse. Pain 130(1–2): 108–118.



Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BUM-10.23760005-9366-19055-Hohlbaum.pdf (0.31 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BUM-10.23760005-9366-19055-Hohlbaum_Supplementary.pdf (0.16 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23760005-9366-19055-Hohlbaum_Tabelle 1.pdf (0.12 MB) herunterladen möchten
Kostenfreier Download
Klicken Sie hier, wenn Sie das PDF BMTW-10.23760005-9366-19055-Hohlbaum_Tabelle 2.pdf (0.12 MB) herunterladen möchten



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}